|
|
|
Free Neuropathology 5:15 (2024) |
|
Methods |
|
An optimized filter trap assay for detecting recombinant authentic tau fibrils |
|
Allison R. Balaj1, Thomas L. Rothstein1, Hiroaki Kaku1 |
|
|
Corresponding author: |
|
Additional resources and electronic supplementary material: supplementary material |
|
Submitted: 23 April 2024 |
|
Keywords: Truncated tau fibrils, Filter trap assay, Sarkosyl, Tauopathies, Protein aggregation |
|
Abstract The development and optimization of the Filter Trap Assay (FTA) for the detection of authentic tau fibrils in vitro mark a pivotal advancement in the realm of tauopathy research, particularly by addressing the limitations of using polyanion-induced tau fibrils, which structurally differ from those isolated from tauopathy patients. Recently it has been shown that truncated tau fragment (297-391), also termed dGAE, can form authentic tau fibrils in the absence of polyanions. This study introduces a refined protocol that reliably detects authentic tau fibrils in a physiologically relevant framework, utilizing nitrocellulose membranes to achieve heightened sensitivity. Our investigation highlights the superior efficacy of sarkosyl, an anionic surfactant traditionally used to prepare protein lysates from brains and cultured neurons, in preserving the aggregated state of tau dGAE fibrils in vitro, underscoring its potential for further exploratory studies. By offering a user-friendly and economically feasible approach, this technique enables a broad range of laboratories to measure the presence of authentic tau fibrils. This methodological enhancement propels our understanding of tauopathies forward and bridges the gap between basic research and advanced structural analyses, enriching the scientific community's methodologies for studying neurodegenerative disorders. |
|
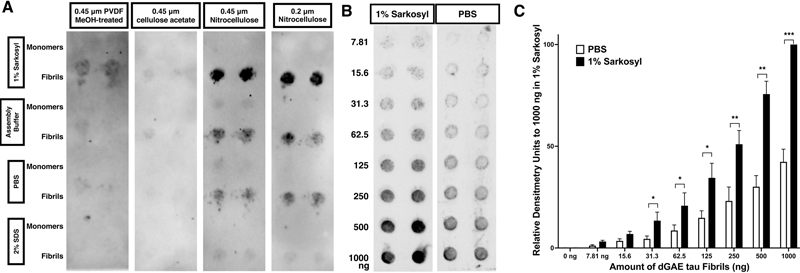
Introduction The microtubule-associated protein tau forms disease-specific filamentous aggregates in tauopathies, including Alzheimer’s diseases (AD). Recent advances in cryo-electron microscopy (cryo-EM) have made it a powerful tool for determining the 3D structure of various proteins, including tau aggregates. The structure of aggregated tau from AD patients was determined: Tau filament cores are made of two identical protofilaments comprising residues 306-378 of Tau, which adopt a combined cross-β/β-helix structure and define the seed for tau aggregation1. To model tau aggregation in vitro, polyanions, such as heparin, have been widely used to mimic tau aggregation2,3. However, the 3D structure of heparin-induced tau fibrils is quite different from those found in patients. Unlike the uniform structure observed in disease-related tau, heparin-induced tau fibrils are a mixture of three distinct structures: snake, twister, and jagged4. Thus, heparin-induced tau aggregates may not represent a physiological model for in vitro studies. This raises important considerations about the relevance and accuracy of using heparin-induced models in tau aggregation research. To overcome this issue, methods to generate tau aggregates without the need for polyanions were identified using the aggregation core region, such as truncated tau 297-391, termed dGAE tau5,6. This breakthrough offers new avenues for studying tau aggregation more physiologically. The filter trap assay (FTA), also known as the filter retardation assay, is a widely recognized, user-friendly in vitro method for detecting protein aggregates including polyanion-induced tau aggregates. Given the structural differences between polyanion-induced tau aggregates and those extracted from tauopathy patients, work on the aggregation of the dGAE tau variant is expected to increase. A common approach to assess dGAE tau aggregation involves the use of Thioflavin T (ThT) fluorescence assays. However, these fluorescence-based assays sometimes yield inconsistent results, necessitating alternative methods to verify findings. In this context, FTA, which utilizes a distinct mechanism for detecting tau aggregates, emerges as an essential complementary technique. While FTA detection methods for polyanion-induced tau aggregates have been previously established, the methods for detecting authentic tau fibrils remain largely unexplored7. The notable structural differences between these fibrils suggest distinct biochemical characteristics. Herein, we present an optimized FTA protocol for analyzing dGAE tau aggregates. This protocol is designed to provide more reliable and reproducible results than previous protocols, thereby enhancing the study of tau aggregation dynamics in vitro and contributing to a deeper understanding of tauopathies. Results Initially, we followed the FTA protocol published in 20087. According to this protocol, tau protein samples are diluted in 2 % (w/v) sodium dodecyl sulfate (SDS) and the FTA is conducted using three types of membranes: nitrocellulose, cellulose acetate, and polyvinylidene fluoride (PVDF)7. Our findings indicated that the nitrocellulose membrane yielded the highest sensitivity. Notably, the sensitivity of dGAE tau fibrils in the FTA diminished when diluted in 2 % SDS (Figure 1A). Subsequently, we explored FTA using various solutions, such as phosphate buffered saline (PBS), 1 % (w/v) sarkosyl, and assembly/HEPES (4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid) buffer, all of which have been previously documented for suspending whole tau fibrils6,8,9. Tau dGAE fibrils maintained their aggregated form more effectively when diluted in 1 % sarkosyl (Figure 1A). No significant difference in sensitivity was observed between nitrocellulose membranes with pore sizes of 0.2 μm and 0.45 μm (Figure 1A). As little as 7.8 ng of dGAE fibrils could be detected using a 0.45 μm pore size nitrocellulose membrane when diluted in 1 % sarkosyl (Figures 1B and 1C).
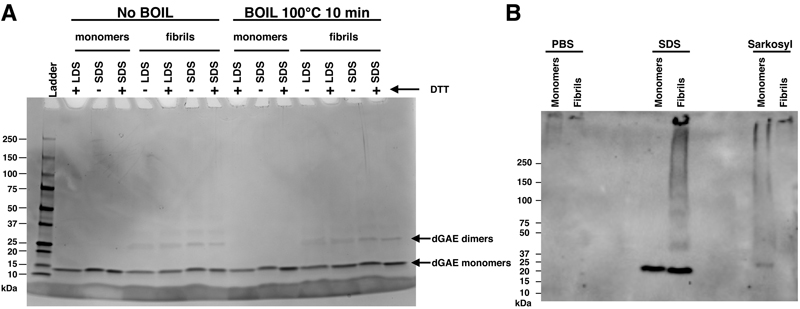
Figure 1. Optimal sensitivity in detecting dGAE tau fibrils is achieved using a 1 % sarkosyl solution in conjunction with a nitrocellulose membrane Using SDS-PAGE we found that the majority of dGAE tau aggregates were reduced to monomers when treated with Laemmli sample buffers containing SDS or lithium dodecyl phosphate (LDS), regardless of heat denaturation and reducing processes (Figure 2A). To corroborate our observations, we subjected protein samples, prepared in PBS, SDS, and sarkosyl, to native-gel PAGE followed by western blotting. Both the monomeric and fibrillar forms of tau dGAE protein, when in PBS, remained in the wells, presumably due to the high basic nature of tau dGAE, which has a theoretical isoelectric point of 9.59 and thus would be positively charged in the absence of an anionic surfactant10. When subjected to anionic detergents, the dGAE tau proteins displayed distinct behaviors: in SDS, the fibrils predominantly were broken down into monomers, whereas in sarkosyl, they retained their fibrillar integrity (Figure 2B).
Figure 2. The majority of dGAE tau aggregates are reduced to monomers when treated with SDS Taken together, these data imply that for effective FTA detection, tau dGAE samples should be diluted in sarkosyl that preserves their aggregated state. Discussion The introduction of truncated tau variants such as dGAE tau offers a more representative in vitro model for studying tau aggregation as compared polyanion tau aggregation. We optimized a protocol for the FTA using dGAE tau aggregates, representing a notable stride towards more accurate and physiologically relevant in vitro models. The use of nitrocellulose membranes in the FTA emerged as a key factor in achieving high sensitivity in detecting tau aggregates, thereby providing a robust alternative to ThT fluorescence assays, which are prone to inconsistency. Moreover, our exploration of different solvents such as PBS, 1 % sarkosyl, and assembly/HEPES buffer for FTA further refines the approach to study tau aggregation. Although sarkosyl has traditionally been used to prepare lysates from brain tissues and from cultured neurons to detect tau aggregates, the effectiveness of sarkosyl in maintaining the aggregated state of tau dGAE fibrils in vitro presents a promising avenue for future studies. The underlying mechanism for the enhanced efficiency of sarkosyl in detecting tau dGAE aggregates through FTA compared to PBS and assembly/HEPES buffer remains elusive. We speculate that the unique physicochemical properties of sarkosyl may interact with tau aggregates in a manner that is less denaturing than SDS, or promotes the stabilization of tau aggregates, thus facilitating their capture on the membrane. Further investigations are required to delineate the specific molecular interactions and biophysical changes induced by sarkosyl, which may offer insights into its preferential efficacy and may potentially uncover novel aspects of tau aggregation dynamics. Thus, the assay we developed in this study is especially relevant given the need for more reliable and reproducible methods in tauopathy research, particularly in view of its simple, efficient, user-friendly and cost-effective approach. This ease of use and affordability make FTA an accessible tool for laboratories with varying resources, democratizing research in neurodegenerative diseases. This method offers a practical alternative to more complex and expensive techniques, such as EM. While cryo-EM provides unparalleled detail in structural analysis, its high cost and technical requirements can be prohibitive. In contrast, the FTA protocol, especially with the use of nitrocellulose membranes, presents a straightforward and economical solution for initial screening and analysis of authentic tau aggregates. This approach not only facilitates routine laboratory investigations but may also serve as a critical preliminary screening step before employing more sophisticated methods. The capacity to detect minimal quantities of dGAE fibrils (as low as 7.8 ng) using affordable materials underscores its practicality for widespread application. The flexibility of the method, demonstrated by the effective use of different solvents like sarkosyl, adds to its appeal in diverse research settings. This study therefore positions the optimized FTA protocol as an excellent tool in the arsenal of in vitro cell-free tauopathy research, enabling more laboratories to engage in meaningful investigation of tau aggregation. It paves the way for the use of more intricate techniques such as EM to further elucidate the complex mechanisms underlying tauopathies. By providing a cost-effective and user-friendly method, we bridge the gap between basic research and advanced structural analysis, fostering a more comprehensive understanding of tauopathies. Author Contributions Conceptualization, H.K.; software, H.K.; investigation, A.R.B. and H.K.; resources, H.K.; data curation, A.R.B. and H.K.; writing - original draft preparation, H.K.; writing - review and editing, A.R.B., T.L.R. and H.K.; visualization, H.K.; supervision, H.K.; project administration, H.K.; funding acquisition, T.L.R. and H.K. Acknowledgments We are grateful to our colleagues for helpful discussions and technical assistance throughout the course of this study. This work was supported by the National Institute on Aging of the National Institutes of Health (NIH) under Award Number R03 AG072148. Data Availability Statement The datasets generated and/or analyzed during the current study are available on demand. Conflicts of Interest Statement The authors declare no conflicts of interest. Funding Statement This work was supported by the Pilot Research Project grant awarded by the Western Michigan University Homer Stryker M.D. School of Medicine, and Public Health Service grant AG072148 awarded by the National Institutes of Health. References 1. Fitzpatrick, A. W. P. et al. Cryo-EM structures of tau filaments from Alzheimer's disease. Nature 547, 185-190, https://doi.org/10.1038/nature23002 (2017). 2. Friedhoff, P., Schneider, A., Mandelkow, E. M. & Mandelkow, E. Rapid assembly of Alzheimer-like paired helical filaments from microtubule-associated protein tau monitored by fluorescence in solution. Biochemistry 37, 10223-10230, https://doi.org/10.1021/bi980537d (1998). 3. Montgomery, K. M. et al. Chemical Features of Polyanions Modulate Tau Aggregation and Conformational States. J Am Chem Soc 145, 3926-3936, https://doi.org/10.1021/jacs.2c08004 (2023). 4. Zhang, W. et al. Heparin-induced tau filaments are polymorphic and differ from those in Alzheimer's and Pick's diseases. eLife 8, https://doi.org/10.7554/eLife.43584 (2019). 5. Al-Hilaly, Y. K. et al. Tau (297-391) forms filaments that structurally mimic the core of paired helical filaments in Alzheimer's disease brain. FEBS Lett 594, 944-950, https://doi.org/10.1002/1873-3468.13675 (2020). 6. Lovestam, S. et al. Assembly of recombinant tau into filaments identical to those of Alzheimer's disease and chronic traumatic encephalopathy. eLife 11, https://doi.org/10.7554/eLife.76494 (2022). 7. Chang, E. & Kuret, J. Detection and quantification of tau aggregation using a membrane filter assay. Anal Biochem 373, 330-336, https://doi.org/10.1016/j.ab.2007.09.015 (2008). 8. Greenberg, S. G. & Davies, P. A preparation of Alzheimer paired helical filaments that displays distinct tau proteins by polyacrylamide gel electrophoresis. Proc Natl Acad Sci U S A 87, 5827-5831, https://doi.org/10.1073/pnas.87.15.5827 (1990). 9. DeSantis, M. E. et al. Operational plasticity enables hsp104 to disaggregate diverse amyloid and nonamyloid clients. Cell 151, 778-793, https://doi.org/10.1016/j.cell.2012.09.038 (2012). 10. Gasteiger, E. et al. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res 31, 3784-3788, https://doi.org/10.1093/nar/gkg563 (2003).
Copyright: © 2024 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |