|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Free Neuropathology 5:3 (2024) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Original Paper |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cellular activation patterns of CD10+ fibro-adipogenic progenitors across acquired disease states in human skeletal muscle biopsies |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Peter W. Schutz1, Simon Cheung2, Lin Yi3, Fabio M. V. Rossi3 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Corresponding author: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Submitted: 01 November 2023 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Keywords: Fibro-adipogenic progenitor, Inflammatory myopathy, Denervation, CD10, Periarteriolar, Type 2 atrophy |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Abstract
Background: Fibro-adipogenic progenitors (FAP) are muscle resident mesenchymal stem cells pivotal for regulation of myofiber repair. Experimental results show in addition involvement in a range of other pathological conditions and potential for pharmacological intervention. FAP histopathology in human muscle biopsies is largely unknown, but has potential to inform translational research. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Introduction Fibro-adipogenic progenitor cells (FAPs) are muscle resident mesenchymal stem cells with potential to differentiate along a variety of lineages (fibrogenic, adipogenic, osteogenic) as well as several defined functions in muscle regeneration and tissue homeostasis. Elegant mouse studies have conclusively outlined their central role in determining muscle fiber repair or fibrosis after acute injury via cellular interactions with myogenic muscle stem cells (satellite cells) and macrophages [18]. Subsequent studies have demonstrated their involvement in mediating neurogenic atrophy, highlighting involvement in two very different types of tissue response [9, 20]. More specific pathomechanistic roles have been demonstrated experimentally in the context of muscular dystrophies (dystrophinopathies; dysferlinopathy, facioscapulohumeral muscular dystrophy, fibrodysplasia ossificans progressiva, posttraumatic fibroadipogenic muscle degeneration (rotator cuff tears), and fibro-fatty muscle degeneration in the context of systemic disease (chronic renal insufficiency, type 2 diabetes), and senescent cell states in inflammatory myopathies [5, 6, 14, 16-19, 21, 22, 25]. Quiescent FAPs are present in endomysial, perimysial, epimysial and perivascular locations [3]. Within the endomysium, FAPs are situated in the interstitial space outside basement membranes. Endomysial FAPs form an interconnected network of processes on the surface of muscle fibers and extending along capillaries. CD10 is a robust and widely available marker for FAPs in human muscle [12]. In response to activating signals, FAPs can initiate intercellular communications, clonal expansion, differentiation, or subsequent apoptosis [3, 30]. Morphological changes in response to activation in human skeletal muscle are not well delineated. While the involvement of FAPs in various pathogenic contexts is coming to light, single-cell RNA sequencing has uncovered transcriptomic heterogeneity of molecular FAP subpopulations in animal models and in humans [7, 13, 15, 21]. Recent evidence indicates, for instance, that a CD90+ FAP subpopulation increases selectively in skeletal muscle of patients with type 2 diabetes [6]. In particular, CD10 has been recently reported to identify a subset of these cells with high adipogenic potential [7]. These data suggest that quiescent FAPs can evolve into a variety of transcriptomic subtypes in the context of a given pathological condition, whose exact biological significance and potential for therapeutic intervention often remain to be elucidated. Experimental proof of concept studies have shown that targeting FAPs pharmacologically has therapeutic potential. Direct interruption of specific FAP signaling pathways with tyrosine kinase inhibitors can reduce fibrosis in a dystrophinopathy mouse model [18], indirect modulation of the muscular cytokine environment can result in FAP mediated augmentation of muscle regeneration [4], and the beneficial effects of exercise on senescent muscle are partly mediated by FAPs [25]. In view of these results, understanding pathogenetic contributions of FAPs in a wider range of human myopathies is of translational interest. Little is known about the histological appearance of activated FAPs in human skeletal muscle biopsies across various myopathies. The majority of FAP studies have been performed in well controlled mouse models, mostly with changes in FAP cell numbers as a measure of activation [17, 18, 20, 21, 25]. Better understanding of histopathological reaction patterns of FAPs on muscle biopsy may help to screen a wide range of conditions for potentially pathological FAP activation. The aim of this study was to delineate histological activation patterns of FAPs in human skeletal muscle biopsies to allow for immunohistochemical screening and evaluation of FAP activation in various conditions. We compared histopathological changes of CD10+ FAPs in six frequent disease conditions seen in clinical muscle biopsies. Methods The study was approved by the Clinical Research Ethics Board of the University of British Columbia. Chart review and biopsy selection Muscle biopsies taken between 2010 and 2018 were selected from the archive of the Muscle Pathology Laboratory at Vancouver General Hospital for nine groups: Normal (N), anti-synthetase syndrome associated myositis (ASA-M), dermatomyositis (DM), inclusion body myositis (IBM), immune mediated necrotizing myopathy (IMNM), type 2 fiber atrophy (T2A), neurogenic atrophy (NA), and rhabdomyolysis (RM). Pathological categories were selected to represent histopathologically well defined acquired myopathic conditions. Electronically available medical records were reviewed. Normal biopsies were chosen based on the absence of abnormalities on routine clinical biopsy evaluation and chart review negative for muscular conditions. Histology and chromogen immunohistochemistry Paraffin sections (4 µm) were used for evaluation in order to take advantage of established CD10 immunohistochemical protocols. HE stained sections were prepared in the usual manner. CD10 was used as marker for FAPs based on poor performance of PDGFR-α immunohistochemistry in human muscle samples and previous work by Hejbol et al lending support to the use of CD10 as FAP marker in human muscle tissue [12]. All immunohistochemical stains were performed on a Dako Omnis instrument with heat epitope retrieval for 30 min at 97C and pH 9.0. The CD10 antibody used was Dako ready to use monoclonal, clone 56C6 (Agilent, Santa Clara, USA), with DAB chromogen. Fluorescence immunohistochemistry Normal human muscle frozen section (8 µm) was fixed in cold Acetone/Ethanol (1:1) for 10 min, followed by washing in PBS for 30 min. After blocking in 20% normal goat serum for 30 min, sections were concomitantly incubated overnight at 4°C with CD10 antibody, clone 56C6 (Bio-Rad, MCA1806T, isotype mouse IgG1, dilution 1:40), and Laminin-2-alpha antibody, clone 4H8-2 (Abcam, ab11576, isotype rat IgG1, dilution: 1:200). After washing, section was stained for 2 hrs. at room temperature with secondary antibodies Goat anti-Mouse IgG1 Alexa Fluor 647 (Thermo Fisher, 21240, dilution 1:1000) and Goat anti-Rat IgG Alexa Fluor 488 (Thermo Fisher, A-11006, dilution 1:1000). Hoechst 33342 was used for nuclear staining. Histopathological review Stained sections were reviewed by an experienced myopathologist (PWS). Qualitative changes were captured descriptively on initial review. The type of qualitative changes found was suggestive of a semi-quantitative scale to compare changes between biopsies (Table 2. Figure 4.). This semi-quantitative scale could be consistently applied to all biopsies. The scoring system is entirely descriptive at this stage and any potential pathological significance could be addressed in subsequent studies. Results Patient characteristics We reviewed muscle biopsies from 32 patients summarized in table 1. The age range was 39 – 82 years, biopsied muscles included quadriceps, biceps, gastrocnemius, and, in a single case, deltoid. Gastrocnemius biopsies are over-represented in the neurogenic atrophy group since these biopsies were often taken concomitantly with sural nerve biopsies. There is a slight male predominance in the IBM group in keeping with disease epidemiology. The selection of normal controls was based on normal myopathological examination and absence of neuromuscular disease upon retrospective chart review. Clinical indications for biopsies with normal muscle histology are listed. Serological information for patients with inflammatory myopathies was available in 6/9 cases and is presented in table 1. Diagnosis of inflammatory myopathies in cases without serological information is based on clinico-pathological correlation.
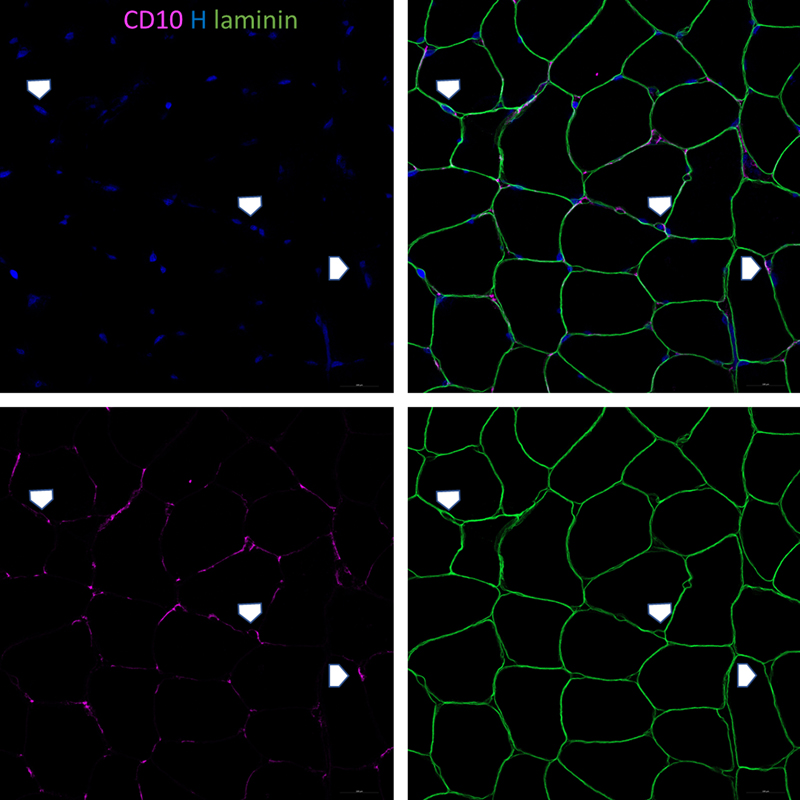
Table 1. Clinical and serological characteristics of study cases. N: Normal; ASA-M: Anti-synthetase autoantibody associated myositis; DM: Dermatomyositis; IBM: Inclusion body myositis; IMNM: Immune mediated necrotizing myopathy; T2A: Type 2 fiber atrophy; NA: Neurogenic atrophy; RM: Rhabdomyolysis; MSA: Myositis-specific-antibody; Q: Quadriceps femoris; B: Biceps brachii; D: Deltoid; G: Gastrocnemius. CD10+ FAPs in normal skeletal muscle biopsies The histological appearance of FAPs revealed by CD10 immunohistochemistry was consistent with expectations based on previous studies [12]. Cross sections of skeletal muscle showed scattered concave triangular or quadrangular cell bodies in endomysial corners between muscle fibers (Fig. 1A). Linear endomysial spaces between muscle fibers show occasional wispy lines of CD10 positivity, corresponding FAP cell processes running along the surface of muscle fibers in a transverse plane. These processes are clearly visible on longitudinal sections tangential to the surface of muscle fibers, which show delicate branching processes embracing muscle fibers (Fig. 1B). Longitudinal sections through the body of muscle fibers demonstrate dot-like staining in the endomysial space, representing delicate processes in cross section. There is band-like or coarsely granular staining along capillaries, which likely reflects the known proclivity of FAPs to arrange along capillaries. Small vessels with thicker walls, likely arteriolar, show delicate sparse perivascular staining (Fig. 1G). Localization of CD10+ FAPs outside basement membrane was confirmed on immunofluorescent triple labelling (Fig. 2).
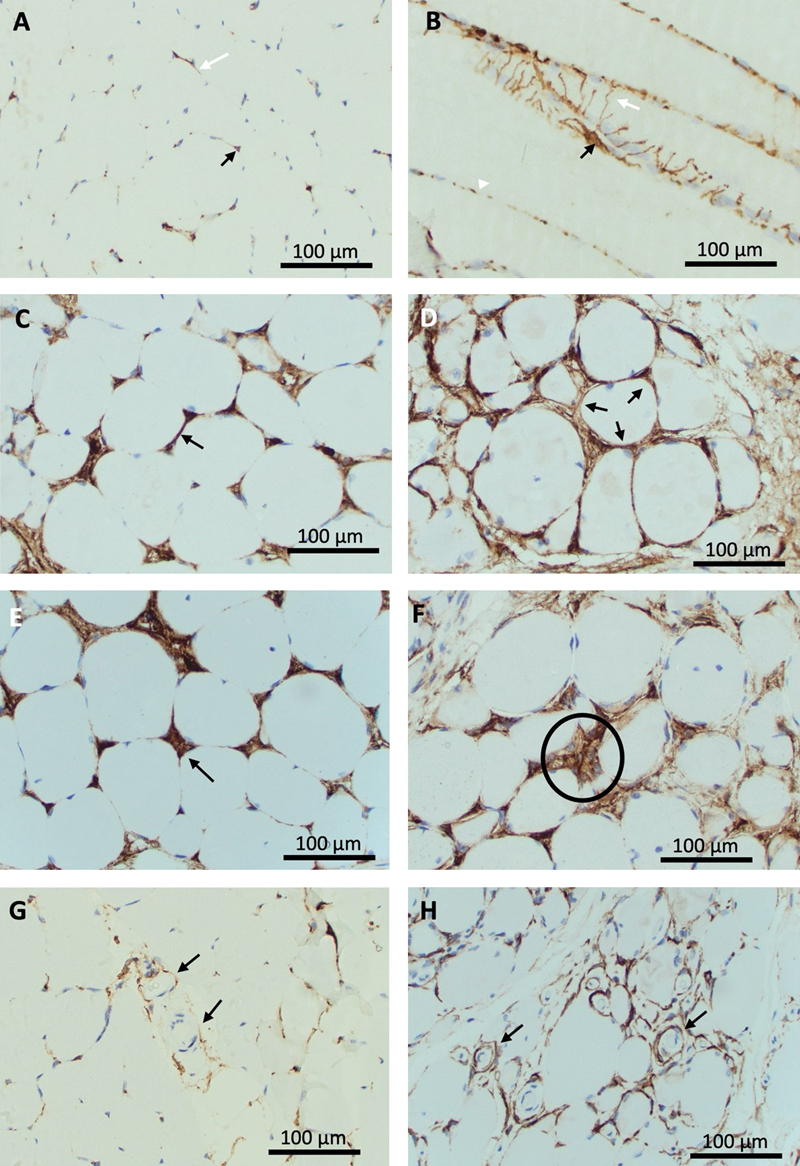
Figure 1. Resting and activated histology of CD10+ fibro-adipogenic progenitors in human skeletal muscle.
A-B: Normal muscle demonstrates occasional triangular staining cell bodies in endomysial corners (black arrow) and delicate linear staining processes along endomysial lines (white arrow) on cross section (A). Longitudinal sections (B) show plump cell bodies (black arrow) with processes extending around muscle fibers (white arrow). These appear as dot-like staining on deeper planes (white arrowhead).
Figure 2. Normal muscle biopsy with immunofluorescent labelling for laminin, CD10, and Hoechst demonstrates location of plump shaped CD10+ cells bodies and wispy processes outside the basal lamina in the endomysial space, in keeping with the expected location of CD10+ FAPs. Cellular changes of CD10+ FAPs in pathological muscle biopsies Qualitative review showed changes to the appearance of cell processes as well as to cell bodies in pathological biopsies. Processes frequently appear as ropey and coarse linear staining along endomysial lines, either segmental or circumferential around muscle fibers, indicative of expanded, hypertrophic FAP process networks (Fig. 1C, D). Longitudinal sections demonstrate broad sheets of CD10 positivity instead of delicate thread-like processes. Endomysial corners can show enlarged, prominent, bulky staining, indicative of expanded or conglomerated cell bodies. Several biopsies show clusters of nuclei in a CD10 positive background, indicating cell clusters as opposed to single, slender angulated cell bodies in normal biopsies (Fig. 1E, F). A subset of biopsies demonstrated strong circumferential staining around small vessels with a thin muscularis, consistent with enhanced periarteriolar cell processes. This contrasts with sparse delicate periarteriolar staining in normal biopsies (Fig. 1G, H). Overall, the observed changes can be placed into three categories: (i) Expansion of FAP processes represented by ropey thickening and circumferential staining along endomysial lines; (ii) Proliferation and hypertrophy of cell bodies in endomysial corners, represented by the presence of cell clusters; (iii) Expansion of periarteriolar processes, indicated by circumferential coarse staining around thicker-walled small vessels. These changes did not occur in unison and each component could be seen in isolation or in conjunction with others. For semi-quantitative evaluation of the cellular activation of FAPs we developed a scoring system based on these observations, capturing process expansion, cell clusters, and periarteriolar staining. The scoring system is summarized in table 2. Exemplary images for semi-quantitative evaluation are presented in Fig. 3.
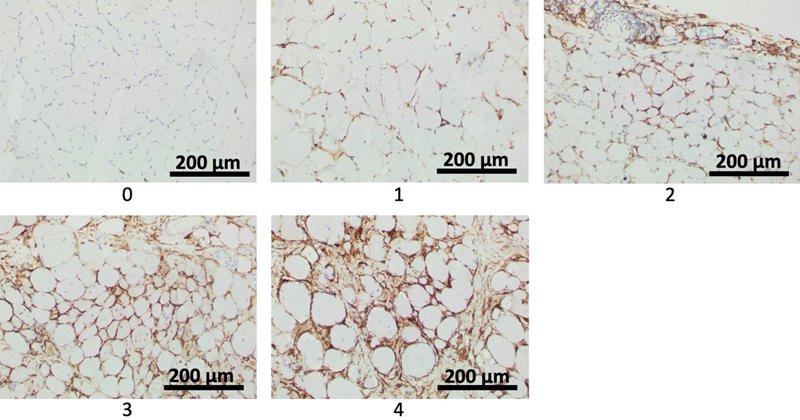
Figure 3. Exemplary photographs representing semiquantitative scores 1 to 4 of endomysial FAP process activation as defined in Table 2. With increasing severity, ropey processes are seen along increasing numbers of myofibers, eventually showing circumferential staining around a few and many fibers for scores 3 and 4, respectively.
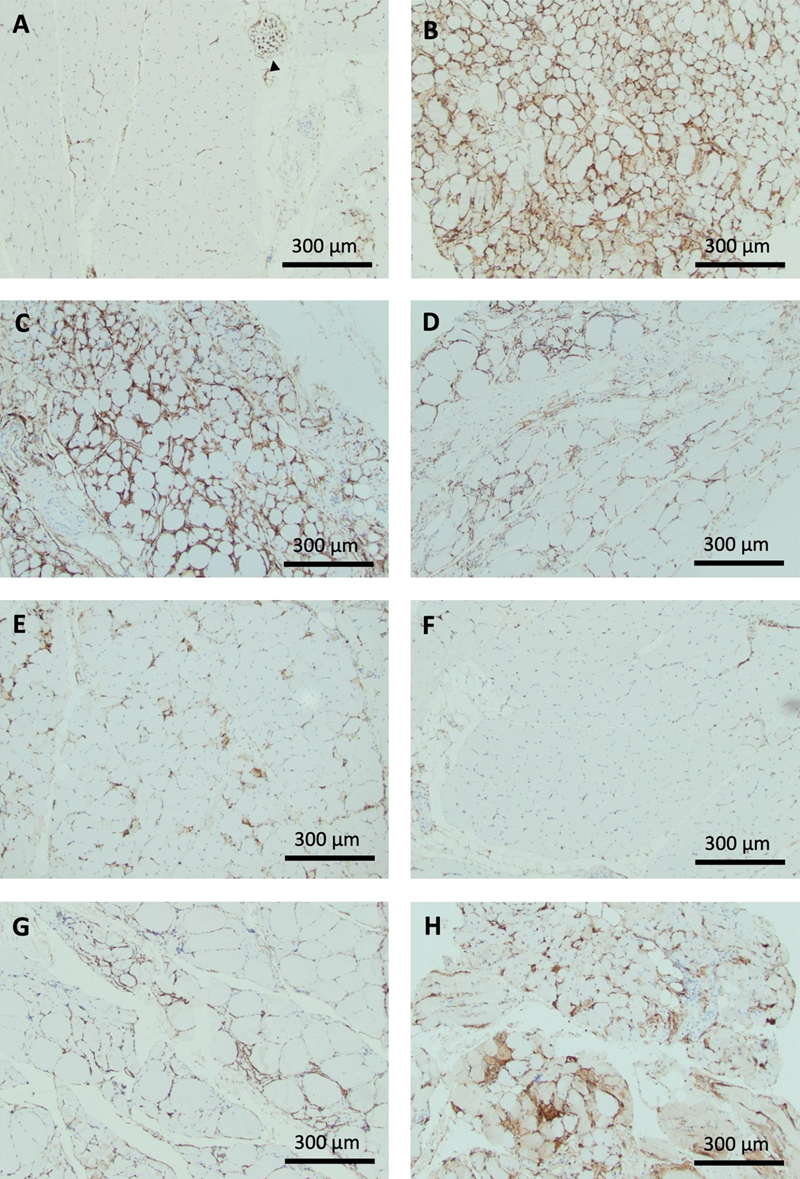
Table 2. Semi-quantitative scoring system for endomysial and periarteriolar FAP activation on CD10 immunohistochemically stained cross sections of skeletal muscle. Endomysial activation involves cell process expansion and/or evidence of proliferation in the form of cell clusters. Periarteriolar activation is represented by arterioles entirely surrounded by CD10+ processes. For illustration of histological changes see figure 1. For exemplary photos of endomysial FAP process activation scores see figure 4. Immunoreactivity for CD10 of non-FAP structures in muscle biopsies CD10 labels FAPs in human muscle biopsies, but it is not a specific marker. Its utility derives from a combination of the characteristic morphology of FAPs with CD10 positivity. Other histologically easily identifiable structures can label for CD10. Sarcoplasm of regenerating fibers can show positivity for CD10 [2], as was apparent in biopsies for IMNM and rhabdomyolysis (Fig. 4E, H). Myelin sheaths and perineurium label with CD10 [12], seen in a small nerve branch in the normal biopsy represented in Fig. 4A. Although CD10 is expressed in early lymphoid progenitors and germinal center cells, these are not normally part of inflammatory infiltrates in muscle, unless histologically distinct lymph-follicle-like-structures are present as part of an inflammatory process, or there is involvement by CD10+ lymphoma.
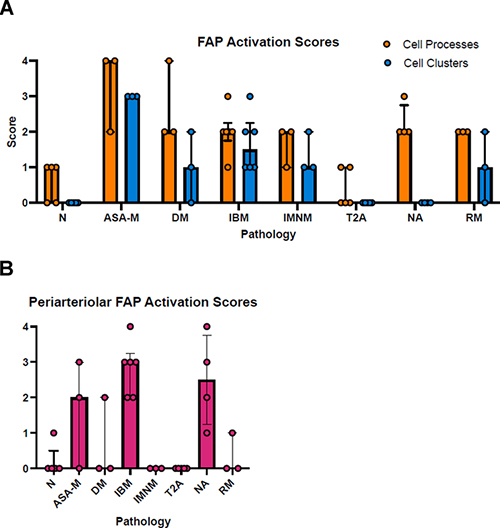
Figure 4. Exemplary patterns of CD10+ FAP activation across pathological groups. A: Normal muscle biopsy. Occasional triangular cell bodies in endomysial corners and granular staining along capillaries running transversely. There is in addition a cross section of peripheral nerve with CD10 immunoreactivity of perineurium and myelin sheaths (black arrowhead). Endomysial and periarteriolar CD10+ FAP activation across pathological groups Patterns of CD10+ FAP activation differed across disease groups. Typical photographs are presented in Fig. 4, scoring results are summarized in Fig. 5. Detailed correlation of scoring results, HE pathology, and distribution of CD10+ FAP activation is presented in table 3. Biopsies from the anti-synthetase syndrome group showed strong diffuse activation of FAPs with often circumferential processes and prominent FAP cell clusters. On HE stained sections, scattered or perifascicular de- and regenerating fibers were apparent, which demonstrated sarcoplasmic labelling on CD 10 immunohistochemistry. FAP activation was diffuse and as such included activation around normal appearing fibers as well as activation adjacent to degenerating and regenerating fibers (Fig. 4B). Dermatomyositis biopsies showed impressive diffuse circumferential ropey staining in regions of fiber atrophy but notably also in areas of fibers without evidence of de- or regeneration. In addition, there were clusters of cell proliferation, though less pronounced than in ASA-M biopsies. The cause of diffuse and pronounced FAP activation was not obvious on this study in the absence of evidence of fiber de- or regeneration, inflammatory infiltrates, or neurogenic change (Fig. 4C). IBM biopsies showed staining associated with inflammatory foci or regions of chronic myopathic change and fibrosis only, often with cell clusters. This could be interpreted as in keeping with a role of FAPs in fiber repair and fibrosis (Fig. 4D). Interestingly, IBM biopsies tended to show prominent periarteriolar staining, the significance of which is uncertain (Fig. 5B). Immune mediated necrotizing myopathies showed multifocal increased staining restricted to degenerating/regenerating fibers, with a mix of linear coarseness and expanded endomysial corners, consistent with fiber repair (Fig. 4E). Type 2 fiber atrophy did not show significant abnormal staining and has the same profile on scores as normal biopsies (Figs. 4F; 5A). In contrast, biopsies with neurogenic atrophy clearly exhibited increased ropey staining surrounding muscle fibers without evidence of FAP proliferation, as is also apparent on the scoring profile (Figs. 4G; 5A). Neurogenic biopsies showed in addition significant periarteriolar activation (Fig. 5B). Experimental evidence suggests a causal role of FAPs in neurogenic fiber atrophy. The absence of evidence of FAP proliferation raises the possibility that this effect may primarily go along with cell process expansion in human skeletal muscle. Biopsies from patients with rhabdomyolysis represent a histologically more diverse group with biopsies taken at various stages of degeneration or regeneration and with overall variable severity. These biopsies showed variable interstitial staining centered on abnormal fibers and frequent fibers with sarcoplasmic positivity in keeping with fiber regeneration (Fig. 4H).
Figure 5. A: Endomysial FAP activation scores for cell processes and cell proliferation across pathological groups as defined in Table 2; median with interquartile range; circles represent individual cases.
Table 3. Endomysial and periarteriolar activation scores for each case in correlation with HE pathology and observed distribution of CD10 staining. N: Normal; ASA-M: Anti-synthetase autoantibody associated myositis; DM: Dermatomyositis; IBM: Inclusion body myositis; IMNM: Immune mediated necrotizing myopathy; T2A: Type 2 fiber atrophy; NA: Neurogenic atrophy; RM: Rhabdomyolysis; cm: Chronic myopathic change, in particular fiber size variability and endomysial fibrosis; d: Degenerating/necrotic fibers; r: Regenerating fibers; pf: Perifascicular. Discussion Morphological changes of CD10+ FAPs upon activation across a range of pathological conditions in human skeletal muscle biopsies in our study combine expansion of endomysial cell processes, clustering of cell bodies between muscle fibers, and prominent circumferential periarteriolar cell processes. FAP proliferation in response to myofiber injury is well described in experimental studies [18, 25]. Expansion of FAP processes has received less attention, but is a prominent histological feature in our study of endomysial CD10+ FAP activation in human muscle biopsies. Morphologically, the expanded network of processes surrounding myofibers suggests enhanced FAP-myofiber/basement membrane interactions, including surveillance and signaling. This is in keeping with a general role for multipotent stromal cells as damage sensors and responders, modulating the local cellular environment [27]. Several observations of cellular FAP reactions are in keeping with previously reported results. Normal muscle biopsies showed delicate CD10+ FAPs without features of histological activation or at most segmental ropey processes along occasional fibers [3, 12]. Disease groups with muscle fiber degeneration and regeneration as part of their histopathology (IMNM, ASA-M, RM) revealed FAP activation of processes and some evidence of cell proliferation mostly in proximity of degenerating or regenerating fibers, in keeping with their role in muscle fiber repair [18]. Neurogenic atrophy biopsies demonstrate activation of endomysial FAP processes without evidence of FAP proliferation in regions of atrophy and, interestingly, around hypertrophic fibers in one biopsy. Mouse work has previously suggested a distinct and pivotal role of FAP activation for denervation atrophy involving IL6-STAT3 signaling, possibly reflected in an expanded network of processes [20]. It is thought that FAP activation may be triggered by disintegration of neuromuscular junctions as a result of nerve degeneration. Taken together, our findings confirm the applicability of major preclinical insights to clinical muscle biopsies, and support the validity of our approach. They also suggest that the type of cellular reaction (proliferation and expansion of processes) could be modulated by the pathogenetic context. It is tempting to speculate that some of this variability could be related to transcriptomic plasticity emerging in several studies [7, 13, 15, 21]. Dermatomyositis biopsies reveal extensive, diffusely distributed CD10 staining of endomysial circumferential processes and cell clusters. The overall degree of CD10 staining was to some extent related to the general severity of underlying pathological change in terms of fiber atrophy, size variability and fibrosis on HE stained sections. However, normal appearing fibers and fibers without evidence of necrosis, regeneration, or inflammatory infiltrates showed pronounced adjacent and circumferential FAP activation. This pattern cannot be easily explained by experimentally and clinically delineated roles for known FAP activation in fiber degeneration, neurogenic atrophy, or post-traumatic or systemic fibrofatty degeneration. Rather it suggests extensive pathological FAP activation in dermatomyositis related to an unknown cause. Abnormal IFN1-signaling activation is an important component of the partially understood pathogenesis of dermatomyositis [8, 10, 23], and diffuse distribution of FAP activation could indicate its involvement in the underlying autoimmune process. This would be in keeping with a posited role of FAPs as damage sensors interacting with the immune system [18, 27]. Biopsies of type 2 fiber atrophy showed the same scoring profile as normal biopsies, indicating the absence of FAP activation in this pathology group. This result forms a contrast to obvious FAP activation as part of neurogenic atrophy. Type 2 fiber atrophy is a common pathological reaction pattern in muscle disuse or in various, often metabolic or toxic, myopathic conditions. The absence of FAP activation in type 2 atrophy as opposed to neurogenic atrophy indicates a different level of involvement of interstitial cells in both types of atrophy. Antisynthetase-antibody associated (ASA) myositis shows the highest scores for endomysial FAP activation. This can partly be explained by the presence of degenerating and regenerating fibers. In addition, several regions without obvious fiber necrosis or regeneration demonstrated signs of FAP activation, which is more difficult to understand. ASA-myositis is known to show interferon activation on the analysis of cell homogenates in a different pattern from dermatomyositis, with more prominent interferon 2 and less prominent but active interferon 1 activation [1, 23]. This may be part of an explanation for similarly diffuse FAP activation in antisynthetase syndrome and dermatomyositis. Overall, results point to FAP activation as part of an autoimmune process, perhaps in addition to activation caused by fiber necrosis. Periarteriolar FAP scores revealed variable activation across groups with prominent activation in neurogenic atrophy and IBM biopsies, some activation in dermatomyositis and anti-synthetase syndrome, and no activation in type 2 atrophy and immune mediated necrotizing myopathy. Periarteriolar prominence of FAP processes suggests cellular interaction with perivascular basement membrane, vascular mural cells, or endothelial cells. Experimental evidence supports a specific compartmentalized interaction between FAPs and capillaries mediated by VEGF in a model of ischemic skeletal muscle injury. This interaction mediates capillary remodeling and vascularization [11, 26]. Periarteriolar prominence of FAP processes would be in keeping with the spatial organization of endothelial cell interaction. In addition, the role of vascular mural cells for skeletal muscle tissue homeostasis has been suggested [24]. In this context, periarteriolar FAP processes raise the possibility of mural cell – FAP interaction. While experimental data support the existence of spatially compartmentalized interaction between FAPs and vascular structures, it remains unclear why there is differential periarteriolar FAP activation across groups of more chronic myopathies, for instance prominent periarteriolar activation in IBM and none in IMNM. It is tempting to speculate that a specific patho-mechanistic context plays a role. The use of CD10 as immunohistochemical marker to visualize FAPs in muscle biopsies has the advantage of easy applicability to a wide variety of archival biopsies. The fact that the method works well on formalin-fixed, paraffin embedded tissue, expands the spectrum of immunohistochemical stains for paraffin embedded muscle biopsies [28, 29], with the advantage that these can be used in laboratories outside dedicated muscle pathology centers. Although CD10 is not specific for FAPs, their characteristic histological appearance allows reliable identification of reaction patterns. Other CD10-positive, non-FAP structures in muscle biopsies were easily identified in our study. Recently, single cell sequencing technologies have yielded insights into transcriptomic heterogeneity of FAP subpopulations in response to various pathological stimuli [7, 13, 15, 21]. These studies suggest that FAP activation results in a dynamic profile of FAP subtypes, which varies with pathological context. More specifically, a recent publication demonstrated in this regard that CD10+ FAPs in humans represent a large FAP subpopulation with adipogenic potential [7]. It is therefore possible that our approach does not capture the entire population of cells, which could be identified as FAPs on a transcriptomic level. Bearing these limitations in mind, cellular reaction patterns delineated by CD10 immunohistochemistry remain a valid insight into CD10+ FAP activation in various disease groups, even if it may not capture the entire FAP spectrum. Conclusion In conclusion, CD10 immunohistochemistry is a robust technique to analyze histological reaction patterns of CD10+ FAPs in routine clinical muscle biopsies. Histopathological features of activation of endomysial CD10+ FAPs in human muscle biopsies include prominent expansion of the network of cell processes surrounding muscle fibers as well as endomysial cell clusters as evidence of proliferation. In addition, prominence of periarteriolar processes is a notable feature in some pathologies. A scoring system for endomysial and periarteriolar activation is proposed. Analysis of biopsies from a range of pathological groups confirms cellular reaction patterns in keeping with preclinical insights in many instances. Other observations raise the possibilities of pathological FAP activation in dermatomyositis, interactions between FAPs and vascular mural cells or endothelial cells in particular in the context of denervation and IBM, and absence of FAP activation in type 2 fiber atrophy. Reliable diagnostic applications, pathomechanistical significance and therapeutic potential remain to be elucidated. References 1. Bolko L, Jiang W, Tawara N, Landon-Cardinal O, Anquetil C, Benveniste O, Allenbach Y (2021) The role of interferons type I, II and III in myositis: A review. Brain Pathology 31(3): e12955. https://doi.org/10.1111/bpa.12955 2. Broccolini A, Gidaro T, Morosetti R, Gliubizzi C, Servidei T, Pescatori M, Tonali PA, Ricci E, Mirabella M (2006) Neprilysin participates in skeletal muscle regeneration and is accumulated in abnormal muscle fibres of inclusion body myositis. J Neurochem 96: 777-789. https://doi.org/10.1111/j.1471-4159.2005.03584.x 3. Contreras O, Rossi FMV, Theret M (2021) Origins, potency, and heterogeneity of skeletal muscle fibro-adipogenic progenitors-time for new definitions. Skelet Muscle 11: 16. https://doi.org/10.1186/s13395-021-00265-6 4. Davies MR, Liu X, Lee L, Laron D, Ning AY, Kim HT, Feeley BT (2016) TGF-beta Small Molecule Inhibitor SB431542 Reduces Rotator Cuff Muscle Fibrosis and Fatty Infiltration By Promoting Fibro/Adipogenic Progenitor Apoptosis. PLoS One 11: e0155486. https://doi.org/10.1371/journal.pone.0155486 5. Dong J, Dong Y, Chen Z, Mitch WE, Zhang L (2017) The pathway to muscle fibrosis depends on myostatin stimulating the differentiation of fibro/adipogenic progenitor cells in chronic kidney disease. Kidney Int 91: 119-128. https://doi.org/10.1016/j.kint.2016.07.029 6. Farup J, Just J, de Paoli F, Lin L, Jensen JB, Billeskov T, Roman IS, Comert C, Moller AB, Madaro L et al (2021) Human skeletal muscle CD90(+) fibro-adipogenic progenitors are associated with muscle degeneration in type 2 diabetic patients. Cell Metab 33: 2201-2214 e2211. https://doi.org/10.1016/j.cmet.2021.10.001 7. Fitzgerald G, Turiel G, Gorski T, Soro-Arnaiz I, Zhang J, Casartelli NC, Masschelein E, Maffiuletti NA, Sutter R, Leunig M et al (2023) MME+ fibro-adipogenic progenitors are the dominant adipogenic population during fatty infiltration in human skeletal muscle. Commun Biol 6: 111. https://doi.org/10.1038/s42003-023-04504-y 8. Gallay L, Fermon C, Lessard L, Weiss-Gayet M, Viel S, Streichenberger N, Corpet A, Mounier R, Gitiaux C, Mouchiroud G et al (2022) Involvement of Type I Interferon Signaling in Muscle Stem Cell Proliferation During Dermatomyositis. Neurology 98: e2108-e2119. https://doi.org/10.1212/WNL.0000000000200271 9. Gonzalez D, Contreras O, Rebolledo DL, Espinoza JP, van Zundert B, Brandan E (2017) ALS skeletal muscle shows enhanced TGF-beta signaling, fibrosis and induction of fibro/adipogenic progenitor markers. PLoS One 12: e0177649. https://doi.org/10.1371/journal.pone.0177649 10. Greenberg SA, Pinkus JL, Pinkus GS, Burleson T, Sanoudou D, Tawil R, Barohn RJ, Saperstein DS, Briemberg HR, Ericsson M et al (2005) Interferon-alpha/beta-mediated innate immune mechanisms in dermatomyositis. Ann Neurol 57: 664-678. https://doi.org/10.1002/ana.20464 11. Groppa E, Martini P, Derakhshan N, Theret M, Ritso M, Tung LW, Wang YX, Soliman H, Hamer MS, Stankiewicz L et al (2023) Spatial compartmentalization of signaling imparts source-specific functions on secreted factors. Cell Rep 42: 112051. https://doi.org/10.1016/j.celrep.2023.112051 12. Hejbøl EK, Hajjaj MA, Nielsen O, Schrøder HD (2019) Marker Expression of Interstitial Cells in Human Skeletal Muscle: An Immunohistochemical Study. Journal of Histochemistry & Cytochemistry 67: 825-844. https://doi.org/10.1369/0022155419871033 13. Hepler C, Shan B, Zhang Q, Henry GH, Shao M, Vishvanath L, Ghaben AL, Mobley AB, Strand D, Hon GC et al (2018) Identification of functionally distinct fibro-inflammatory and adipogenic stromal subpopulations in visceral adipose tissue of adult mice. Elife 7: e39636. https://doi.org/10.7554/eLife.39636 14. Hogarth MW, Defour A, Lazarski C, Gallardo E, Diaz Manera J, Partridge TA, Nagaraju K, Jaiswal JK (2019) Fibroadipogenic progenitors are responsible for muscle loss in limb girdle muscular dystrophy 2B. Nat Commun 10: 2430. https://doi.org/10.1038/s41467-019-10438-z 15. Jensen JB, Møller AB, Just J, Mose M, de Paoli FV, Billeskov TB, Fred RG, Pers TH, Pedersen SB, Petersen KK et al (2021) Isolation and characterization of muscle stem cells, fibro-adipogenic progenitors, and macrophages from human skeletal muscle biopsies. Am J Physiol Cell Physiol 321: C257-C268. https://doi.org/10.1152/ajpcell.00127.2021 16. Lee C, Agha O, Liu M, Davies M, Bertoy L, Kim HT, Liu X, Feeley BT (2020) Rotator Cuff Fibro-Adipogenic Progenitors Demonstrate Highest Concentration, Proliferative Capacity, and Adipogenic Potential Across Muscle Groups. J Orthop Res 38: 1113-1121. https://doi.org/10.1002/jor.24550 17. Lees-Shepard JB, Yamamoto M, Biswas AA, Stoessel SJ, Nicholas SE, Cogswell CA, Devarakonda PM, Schneider MJ, Jr., Cummins SM, Legendre NP et al (2018) Activin-dependent signaling in fibro/adipogenic progenitors causes fibrodysplasia ossificans progressiva. Nat Commun 9: 471. https://doi.org/10.1038/s41467-018-02872-2 18. Lemos DR, Babaeijandaghi F, Low M, Chang CK, Lee ST, Fiore D, Zhang RH, Natarajan A, Nedospasov SA, Rossi FM (2015) Nilotinib reduces muscle fibrosis in chronic muscle injury by promoting TNF-mediated apoptosis of fibro/adipogenic progenitors. Nat Med 21: 786-794. https://doi.org/10.1038/nm.3869 19. Liu X, Liu M, Lee L, Davies M, Wang Z, Kim H, Feeley BT (2021) Trichostatin A regulates fibro/adipogenic progenitor adipogenesis epigenetically and reduces rotator cuff muscle fatty infiltration. J Orthop Res 39: 1452-1462. https://doi.org/10.1002/jor.24865 20. Madaro L, Passafaro M, Sala D, Etxaniz U, Lugarini F, Proietti D, Alfonsi MV, Nicoletti C, Gatto S, De Bardi M et al (2018) Denervation-activated STAT3-IL-6 signalling in fibro-adipogenic progenitors promotes myofibres atrophy and fibrosis. Nat Cell Biol 20: 917-927. https://doi.org/10.1038/s41556-018-0151-y 21. Malecova B, Gatto S, Etxaniz U, Passafaro M, Cortez A, Nicoletti C, Giordani L, Torcinaro A, De Bardi M, Bicciato S et al (2018) Dynamics of cellular states of fibro-adipogenic progenitors during myogenesis and muscular dystrophy. Nat Commun 9: 3670. https://doi.org/10.1038/s41467-018-06068-6 22. Nelke C, Schroeter CB, Theissen L, Preusse C, Pawlitzki M, Räuber S, Dobelmann V, Cengiz D, Kleefeld F, Roos A et al (2023) Senescent fibro-adipogenic progenitors are potential drivers of pathology in inclusion body myositis. Acta Neuropathol 146(5): 725-745. https://doi.org/10.1007/s00401-023-02637-2 23. Pinal-Fernandez I, Casal-Dominguez M, Derfoul A, Pak K, Plotz P, Miller FW, Milisenda JC, Grau-Junyent JM, Selva-O'Callaghan A, Paik J et al (2019) Identification of distinctive interferon gene signatures in different types of myositis. Neurology 93: e1193-e1204. https://doi.org/10.1212/WNL.0000000000008128 24. Ritso M, Tung LW, Rossi FMV (2022) Emerging skeletal muscle stromal cell diversity: Functional divergence in fibro/adipogenic progenitor and mural cell populations. Exp Cell Res 410: 112947. https://doi.org/10.1016/j.yexcr.2021.112947 25. Saito Y, Chikenji TS, Matsumura T, Nakano M, Fujimiya M (2020) Exercise enhances skeletal muscle regeneration by promoting senescence in fibro-adipogenic progenitors. Nat Commun 11: 889. https://doi.org/10.1038/s41467-020-14734-x 26. Santini MP, Malide D, Hoffman G, Pandey G, D’Escamard V, Nomura-Kitabayashi A, Rovira I, Kataoka H, Ochando J, Harvey RP et al (2020) Tissue-Resident PDGFRα+ Progenitor Cells Contribute to Fibrosis versus Healing in a Context- and Spatiotemporally Dependent Manner. Cell Reports 30: 555-570.e557. https://doi.org/10.1016/j.celrep.2019.12.045 27. Soliman H, Theret M, Scott W, Hill L, Underhill TM, Hinz B, Rossi FMV (2021) Multipotent stromal cells: One name, multiple identities. Cell Stem Cell 28: 1690-1707. https://doi.org/10.1016/j.stem.2021.09.001 28. Suriyonplengsaeng C, Dejthevaporn C, Khongkhatithum C, Sanpapant S, Tubthong N, Pinpradap K, Srinark N, Waisayarat J (2017) Immunohistochemistry of sarcolemmal membrane-associated proteins in formalin-fixed and paraffin-embedded skeletal muscle tissue: a promising tool for the diagnostic evaluation of common muscular dystrophies. Diagnostic Pathology 12(1): 19. https://doi.org/10.1186/s13000-017-0610-y 29. Vogel H, Zamecnik J (2005) Diagnostic Immunohistology of Muscle Diseases. Journal of Neuropathology & Experimental Neurology 64: 181-193. https://doi.org/10.1093/jnen/64.3.181 30. Wei X, Nicoletti C, Puri PL (2021) Fibro-Adipogenic Progenitors: Versatile keepers of skeletal muscle homeostasis, beyond the response to myotrauma. Semin Cell Dev Biol 119: 23-31. https://doi.org/10.1016/j.semcdb.2021.07.013
Copyright: © 2024 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||