|
|
|
Free Neuropathology 1:26 (2020) |
|
Opinion Piece |
|
Enteric synucleinopathy: from trendy concept to real entity. |
|
Adrien de Guilhem de Lataillade 1,2, Thibaud Lebouvier 3, Wendy Noble 4, Laurène Leclair-Visonneau 1, Pascal Derkinderen 1,2 |
|
1 Université de Nantes, Inserm, TENS, The Enteric Nervous System in Gut and Brain Diseases, IMAD, Nantes, France |
|
Corresponding author: |
|
Submitted: 29 July 2020 Accepted: 23 August 2020 Copyedited by: Aivi T. Nguyen Published: 28 August 2020 |
|
Keywords: Synucleinopathies, Tauopathies, Enteric nervous system, Aggregates, Alpha-synuclein, Parkinson’s disease |
|
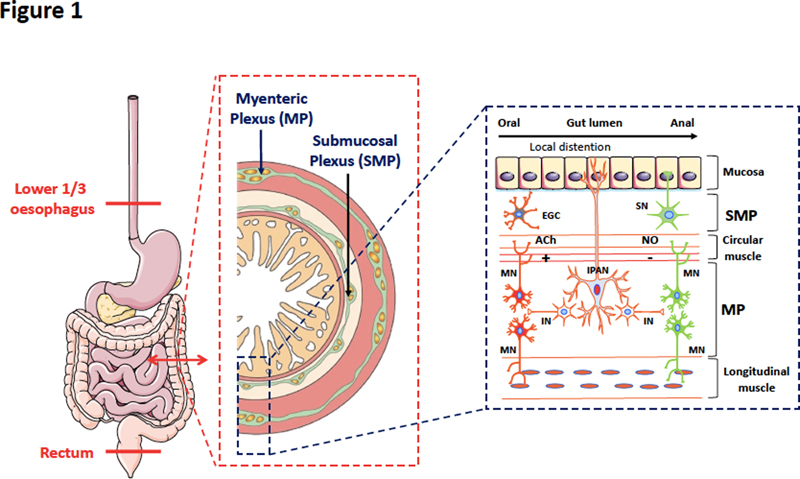
Abstract An accumulating body of literature has emerged in the past 25 years to show that Parkinson’s disease (PD) is not only a disorder of the brain but also of the gastrointestinal tract and more generally of the gut-brain axis. Gastrointestinal symptoms occur in almost every PD patient at some point and in nearly every case examined pathologically autopsy studies find alpha-synuclein deposits, the pathological hallmarks of PD, in the enteric nervous system. This concept of ‘enteric synucleinopathy’ led to the hypothesis that the enteric nervous system might play a pivotal role in the initiation and spreading of PD. Although this hypothesis opens up interesting perspectives on the pathogenesis of neurodegenerative disorders, some important questions are still pending. The present opinion paper describes and compares the physiological and pathophysiological properties of alpha-synuclein in the brain and the enteric nervous system. We conclude that the existing data supports the existence of pathological alpha-synuclein species in the gut in PD. We also discuss if gut-brain interactions are important in other neurodegenerative disorders. Abbreviations AD - Alzheimer’s disease, ENS - enteric nervous system, GI - Gastrointestinal, LB - Lewy bodies, LN - Lewy neurites, PD - Parkinson’s disease, PMCA - protein misfolding cyclic amplification, PSP - progressive supranuclear palsy The enteric nervous system (ENS) is an intricate neural network embedded within the gastrointestinal (GI) tract and distributed from the lower oesophagus to the rectum. Compared to other components of the peripheral nervous system, the ENS shows some unique features that closely resemble the CNS and therefore it is sometimes referred to as ‘the brain-in-the-gut’ or the ‘second brain’ [1] (Figure 1). This close homology between the CNS and ENS suggests that a disease process affecting the CNS could also involve its enteric counterpart. Parkinson’s disease (PD) is the best example of this assumption. PD is the most common synucleinopathy (or synuclein proteinopathy), a group of neurodegenerative disorders characterized by a common pathological lesion composed of aggregates of alpha-synuclein in selectively vulnerable neuron populations in the CNS [2]. Although PD has traditionally been considered a disease of dopaminergic neurons in the substantia nigra, analyses of GI samples from PD patients have consistently found neural pathology, with the presence of alpha-synuclein deposits being detected in the ENS in nearly every PD patient examined [3–11]. This has led to the emerging concept of enteric synucleinopathy. Although these observations open up interesting perspectives on the pathogenesis of neurodegenerative disorders, some important questions are still pending, among which are: what are the biochemical and pathological characteristics of alpha-synuclein deposits in the ENS? Are they similar to those observed in the CNS? Is there any neuronal loss in the ENS in PD? Besides PD, are gut-brain interactions important in other neurodegenerative disorders? Here, we first provide a brief overview of the normal expression profiles of alpha-synuclein in the ENS, before discussing these questions and the arguments for and against the existence of enteric synucleinopathies.
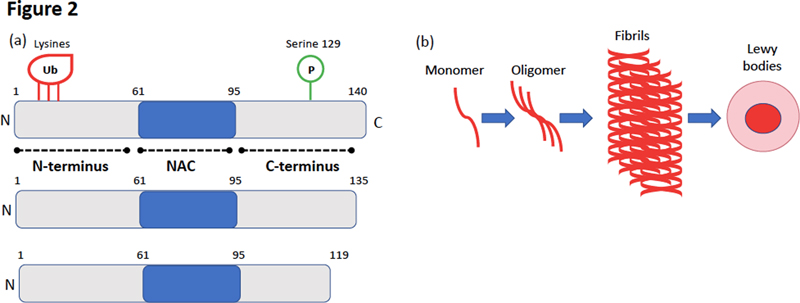
Figure 1. Anatomical organisation and local reflexes of the enteric nervous system. The ENS is a neuronal network embedded within the GI tract and distributed from the lower oesophagus to the rectum. It is organized in two major ganglionated plexuses, the myenteric (MP or Auerbach’s plexus) mainly involved in the control of smooth muscle activity, and the submucosal (SMP or Meissner’s plexus), which regulates secretion (SN, secretory neurons) and microvasculature. Compared to other sections of the peripheral nervous system, the ENS shows unique features that closely resemble some of the CNS: it contains a variety of functionally distinct enteric neurons along with a vast repertoire of neurotransmitters and intercellular messengers which are the basis for enteric neurotransmission. It also harbours a prominent component of glial cells (EGC for enteric glial cells) which, like astrocytes in the CNS, contribute to support, protection and maintenance of the neural networks. Local distention of the intestinal wall and chemical contents in the gut lumen activate intrinsic primary afferent neurons (IPAN) located in both the SMP and MP. The IPAN projects both in oral and anal directions to synapse with interneurons (IN) and motor neurons (MN). Cholinergic (ACh) MN in red are excitatory while nitrergic (NO) MN in green are inhibitory. The peristaltic reflex includes an ascending excitatory reflex mediated by cholinergic MN and elicits contraction of the circular or longitudinal smooth muscles located orally to the site of stimulation. The descending inhibitory reflex involves inhibitory nitrergic MN that elicit relaxation of the circular muscles and longitudinal muscles located anally to the site of stimulation. Alpha-synuclein is physiologically expressed by enteric neurons Alpha-synuclein was first isolated in 1988 from the electric organ of the Pacific electric ray Torpedo californica. In addition to its strong presynaptic localisation, Maroteaux et al., also identified alpha-synuclein in the nucleus, thus accounting for the name ‘synuclein’ (SYNapse + NUCLEus) [12]. Several studies have documented the physiological function of alpha-synuclein in modulating synaptic vesicle release [13]. Human alpha-synuclein, 140 amino acids in length, is mainly expressed by CNS neurons and erythrocytes and is composed of 3 different functional regions (Figure 2a) [13]. There is still open debate about the physiological structure of brain alpha–synuclein as some research groups concluded that it occurs as a helically folded tetramer [14], while others counterclaim that it exists primarily as a disordered monomer [15]. Alpha-synuclein has a natural propensity to aggregate as amyloid structures in a nucleation-dependent process in which monomers assemble via oligomers into fibrils [16] (Figure 2b).
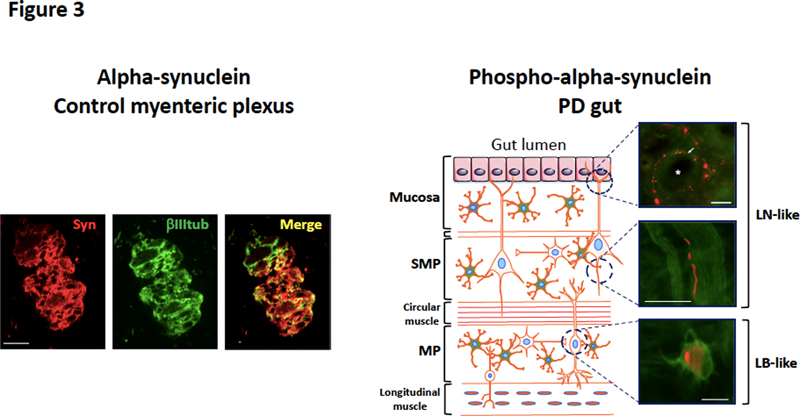
Figure 2. Alpha-synuclein structure and molecular mechanism of its oligomerisation and fibrillogenesis. (a) Schematic representation of alpha-synuclein, which is composed of 3 distinct regions : (i) an N-terminal domain (amino acids 1–60) that binds lipids and contains the lysines that are ubiquitinated (ii) a central domain known as the non-amyloid component (NAC) (amino acids 61–95) which is involved in aggregation, and (iii) a C-terminal acidic tail (amino acids 96–140) accountable for most interactions with other proteins and small molecules and that contains most of the phosphorylation sites including serine 129. Some of the truncated C terminal forms of alpha-synuclein are also shown. (b) Illustration of the molecular steps involved in alpha-synuclein oligomerisation and fibrillogenesis leading to Lewy body formation. Soluble alpha-synuclein is natively unstructured and monomeric. Under pathological conditions, soluble α-synuclein forms β-sheet-like oligomers (protofibrils), which convert into amyloid fibrils and eventually deposit in Lewy bodies. Alpha-synuclein is also expressed by enteric neurons. In rodents, guinea-pigs and human alpha-synuclein positive neurons are present in the two plexuses of the ENS and along the entire digestive tract [5, 17, 18] (Figure 3). Using amine-reactive cross-linking, we showed that, unlike brain neurons, alpha-synuclein exists primarily as a monomer in enteric neurons [19]. As far as we know, the expression of alpha-synuclein by enteric glial cells, the enteric counterpart of CNS astrocytes, has not been evaluated. Detailed immunohistochemical characterisation showed that alpha-synuclein immunoreactive neurons are mostly cholinergic [17, 18] (Figure 1) and that alpha-synuclein is closely associated with the vesicular apparatus [20]. Although the role of alpha-synuclein in the ENS is still mostly unknown, a recent study suggested that it is involved in the development and electrophysiological properties of enteric cholinergic neurons [21].
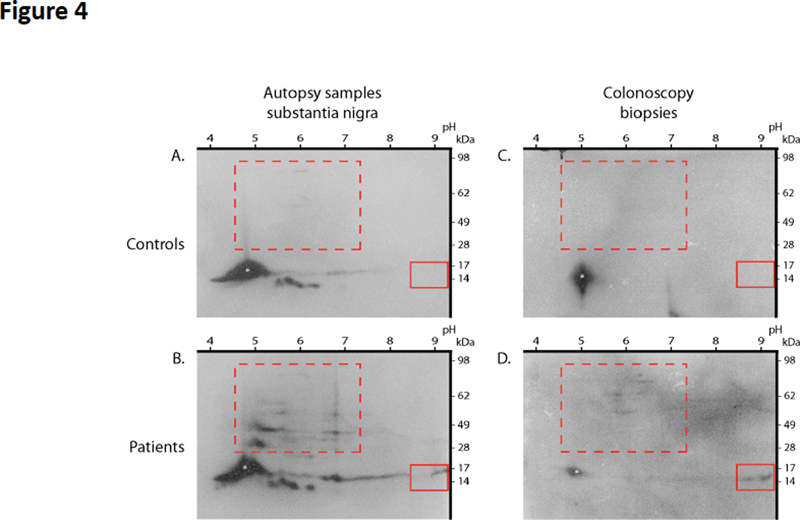
Figure 3. Physiological expression of alpha-synuclein and phospho-alpha-synuclein histopathology in the ENS. Left: anti-alpha-synuclein antibody Syn-1 was used to detect alpha-synuclein in the myenteric ganglia (colon) in a control subject devoid of neurodegenerative disorders; βIII tubulin antibody was used to label the neuronal network. Scale bar is 50 µm. Right: the colon from a PD patient was microdissected into mucosa, whole mount of submucosa and myenteric plexus. Each part was stained with an antibody specific for the phosphorylated form of alpha-synuclein. Phospho-alpha-synuclein histopathology with a LN-like pattern is observed in the mucosa (arrow, the asterisk is for the crypt) and submucosa, while a LB-like structure is found in the myenteric plexus. Scale bar is 40 µm. Does the ENS contain pathological aggregated forms of alpha-synuclein? Lewy bodies (LB) and Lewy neurites (LN) are the defining neuropathological characteristics of PD. LB typically appear in neuronal somata as eosinophilic, rounded inclusions while LN are strand-like structures observed in axons. In 1997, it was demonstrated that LB and LN isolated from PD brain were highly immunoreactive for alpha-synuclein, thereby suggesting that this protein was one of the main components of Lewy pathology [22]. Subsequent work showed that alpha-synuclein in LB and LN was hyperphosphorylated at serine residue 129 [23] (Figure 2). Because alpha and phospho-alpha-synuclein immunohistochemical staining has a much greater sensitivity than hematoxylin and eosin staining for the detection of LB [24], this method quickly became the method of choice for the neuropathological diagnosis of PD [25]. Using this approach, several neuropathology laboratories demonstrated that LB and LN-like structures were observed in the ENS (in both the myenteric and submucosal plexus) in the vast majority of PD patients [3–7] (Figure 3). In some of these studies, proteinase K [3] or alkaline protease [7] pre-treatments were used primarily to unmask antigens and enhance immunolabeling, but such treatments might also allow protease-resistant misfolded, aggregated and hyperphosphorylated alpha-synuclein to be distinguished from soluble forms of the protein. On the whole these findings suggest that pathological aggregated alpha-synuclein is present within the GI tract of PD subjects. Two key issues should however be borne in mind. First, the mere detection of alpha-synuclein phosphorylated at serine 129 is not synonymous with aggregation since soluble alpha-synuclein is also physiologically phosphorylated at this residue [26]. Secondly, immunohistochemical approaches are not sufficient to show that alpha-synuclein is misfolded/aggregated; biochemical confirmation of altered solubility is required. Indeed, in the CNS, a comprehensive biochemical characterisation of alpha-synuclein forms in LB has already been carried out. Using one and two-dimensional immunoblot analysis with modification-specific synuclein antibodies and mass spectroscopy, Anderson et al. confirmed that the predominant modification of alpha-synuclein in LB was phosphorylation at serine 129. They also found a set of additional characteristic modifications including ubiquitination at amino-terminal lysine residues and specific carboxy-terminal truncations [27] (Figure 2). An additional property of alpha-synuclein aggregates found in diseased brains is their seeding ability: the amyloid fibrils formed by alpha-synuclein aggregates can act as templates for the conversion of physiological alpha-synuclein, resulting in the growth of the fibrils and spread of alpha-synuclein pathology [28]. How about the biochemical characterisation of alpha-synuclein in diseased ENS? There are some existing data but it is definitely less complete and robust than those available for the CNS. Using one and two-dimensional analysis of colonic biopsies, we were unable to detect any differences in the expression levels, phosphorylation or aggregation status of alpha-synuclein between controls and PD specimens [29]. These negative findings might however be explained by the relative sparsity of neuronal structures and/or alpha-synuclein inclusions in the GI samples that were used; due to a shortage of samples, only 2 routine colonic biopsies per subject were pooled and analysed. However, when we had the opportunity to analyse 4 pooled biopsies per subject, we were able to detect acidic and high molecular weight alpha-synuclein species in the GI tract of PD patients but not controls, which likely represent hyperphosphorylated and aggregated forms of the protein, respectively (Lebouvier-Derkinderen unpublished results, Figure 4). Such a pattern is reminiscent of that observed in PD brain [27] (Figure 4). The limitations of the two-dimensional immunoblotting technique prompted us to use more sensitive approaches, based on the seeding efficiency of alpha-synuclein. Using an assay inspired by the protein misfolding cyclic amplification (PMCA) assay, we showed that GI biopsies from PD patients (2 to 4 biopsies per subject taken from the upper or lower GI tract) seeded alpha-synuclein aggregation in 10 out of 18 cases [30]. More recently, Viviane Labrie and Patrik Brundin’s group focused on the vermiform appendix, a structure which is particularly enriched in alpha-synuclein [31]. Using mass spectrometry, they identified full-length alpha-synuclein together with a set of 10 truncated forms in the healthy human appendix [32]. Additional experiments performed with an in vitro shaking assay (also inspired by the PMCA approach), showed that appendix lysates from either control or PD subjects seeded aggregation and truncation of alpha-synuclein. These intriguing and provocative findings suggest that aggregated and truncated alpha-synuclein are consistently found in the human healthy appendix, thereby supporting the assumption that the appendix may act as a reservoir for pathogenic forms of alpha-synuclein [31, 32].
Figure 4. High molecular weight and post-translationally modified alpha-synuclein species in the GI tract of PD patients. Immunoblots of total mesencephalon lysates (A and B) from a control (A) and a PD patient autopsy sample (B), and immunoblots of colonic biopsy lysates (C and D) from a control (C) and PD patient (D) were resolved by two-dimensional PAGE using pH 3–10 IEF gradients. Immunoblots were probed with the alpha-synuclein antibody Syn-1. White asterisks mark full-length synuclein. Boxes highlight differences between PD patients and controls: acidic modifications of full-length monomer (solid boxes), and high molecular weight species (dotted boxes) are present in PD patients and absent in controls, in both substantia nigra and colonic biopsies (n=2); pI values and positions of molecular weight standards are indicated. An equivalent amount of protein was loaded in A-B and C-D, respectively. It has also been shown that pathological alpha-synuclein obtained from PD brain has the ability to trigger alpha-synuclein pathology in the CNS of rodents and non-human primates [33, 34]. For example, the intranigral inoculation of alpha-synuclein-containing LB extracts obtained from PD brains leads to widespread alpha-synuclein pathology together with dopaminergic neurodegeneration in mice and monkeys [35]. For the sake of comparisons between brain and enteric alpha-synuclein, it is important to know if pathological alpha-synuclein obtained from the GI tract shares the same pathogenic capacity. Although the precise answer to this question remains unknown, a recent study evaluated the effects of alpha-synuclein aggregates from post-mortem PD stellate ganglia (pooled from 3 patients) injected into mouse brain [36]. Stellate ganglion is a paravertebral ganglion, which like the gut, exhibits marked Lewy pathology in almost all PD subjects [4, 37]. In contrast to the findings obtained with substantia nigra-derived alpha-synuclein, no pathological effects were observed when peripheral aggregates from stellate ganglion were injected into the brain of wild-type mice, at least up to 6 months following injection [36]. This intriguing observation, which casts doubt on the pathogenicity of peripheral alpha-synuclein, requires replication using extracts from other peripheral organs affected by Lewy pathology, such as the GI tract or the salivary glands [3, 38]. Is there any enteric neuronal loss in PD? In the substantia nigra of PD subjects, the presence of alpha-synuclein aggregates is accompanied by severe neuronal loss and clinicopathological findings strongly suggest that the classical PD motor symptoms, including bradykinesia and hypertonia, are driven primarily by neuronal loss rather than only the aggregation of alpha-synuclein [39]. As such, the quantitative evaluation of enteric neuron populations is important to determine the pathological underpinnings of GI symptoms, which are so frequently observed in PD [40]. With that said, and despite the publication of proposed guidelines and consensus techniques, the quantification of submucosal and myenteric neuron number remains challenging [41], mainly because of the fishnet-like architecture of the ENS plexus. In the most comprehensive post-mortem study to date, Annerino et al. used formalin-fixed paraffin embedded sections to compare myenteric neuron density along the length of the GI tract in 6 patients with PD and 12 controls. There were no differences in total myenteric neuron density between controls or patients in any segment examined [42]. Similarly, when whole mounts of submucosa obtained from colonic biopsies were analysed by immunohistochemistry, no major decrease in neuronal density was observed in PD relative to controls in two independent studies that included a total of 58 PD and 30 control cases [8, 43]. On the whole, these data strongly suggest that enteric neuron loss is not a feature of PD. Besides alpha-synuclein, are other aggregated proteins found in the ENS? The observation that alpha-synuclein deposits are a feature of PD gut logically leads to speculation that such a phenomenon might also occur in other neurodegenerative disorders such as tauopathies. Like alpha-synuclein, tau is physiologically expressed by enteric neurons. However, in contrast to the CNS neurons that express all six tau isoforms, adult human ENS primarily express only two tau isoforms [44]. Tau aggregates found in tauopathies generally contain tau in an elevated state of phosphorylation that is often aberrantly cleaved [45]. In a preliminary report, we analysed by Western blot colonic biopsies from 5 patients with probable progressive supranuclear palsy (PSP) using 2 different phospho-tau antibodies and one antibody specific for caspase-cleaved tau [44]. The phosphorylation and truncation patterns of tau in PSP were indistinguishable from those in controls [44]. In a subsequent study, Brittany Dugger et al. performed a comprehensive immunohistochemical analysis of the ENS in tauopathies. Using formalin fixed paraffin embedded sections, they examined the sigmoid colon in 26 PSP, 21 AD and 19 controls using one antibody for total tau and two phospho-tau antibodies [46]. No differences in the staining pattern were observed between colonic specimen from tauopathy patients and controls [46]. So, enteric synucleinopathy: myth or reality? Even if they are still preliminary, recent findings and in particular the ones obtained with ultrasensitive amplification techniques such as PMCA, convincingly showed that alpha-synuclein aggregates with seeding capacity are found in the ENS [30, 32]. It therefore seems justified to use the word ‘enteric synucleinopathy’. That said, some outstanding questions remain: (i) existing findings suggest that enteric neuron loss is not a feature of PD but there could be more subtle morphologic changes [47] that could be evaluated in the future using computer-assisted analysis of the enteric neurons [48] (ii) a comprehensive inventory of alpha-synuclein forms present the ENS from patients with PD still need to be carried out, as has already been performed for the CNS [27] (iii) the results obtained with PMCA need to be independently replicated and confirmed by others amplification techniques such as RT-QuIC (real-time quaking-induced conversion) [49]. In addition, it still remains to be determined if pathological alpha-synuclein purified from the ENS is capable of promoting alpha-synuclein pathology when intracerebrally inoculated, as already demonstrated for CNS-derived alpha-synuclein aggregates [50]. This is a critical issue with regard to PD pathogenesis as it has been speculated that enteric alpha-synuclein aggregates may spread to the CNS via the vagal connections. However, performing such experiments, which require purification of LB-containing fractions from the gut, can be challenging due to the low frequency and density of aggregated alpha-synuclein in the ENS [3, 51]. On the whole, current knowledge strongly supports the existence of enteric synucleinopathy and these data encourage future studies aimed at expanding our knowledge of peripheral pathology in neurodegenerative disorders to gain clues that enable further understanding of the differential pathogenesis of these disorders. References 1. Schemann M, Frieling T, Enck P (2020) To learn, to remember, to forget-How smart is the gut? Acta Physiol (Oxf) 228:e13296. https://doi.org/10.1111/apha.13296 2. Goedert M, Spillantini MG (1998) Lewy body diseases and multiple system atrophy as alpha-synucleinopathies. Mol Psychiatry 3:462–465. https://doi.org/10.1038/sj.mp.4000458 3. Beach TG, Adler CH, Sue LI, et al (2010) Multi-organ distribution of phosphorylated alpha-synuclein histopathology in subjects with Lewy body disorders. Acta Neuropathol 119:689–702. https://doi.org/10.1007/s00401-010-0664-3 4. Gelpi E, Navarro-Otano J, Tolosa E, et al (2014) Multiple organ involvement by alpha-synuclein pathology in Lewy body disorders. Mov Disord 29:1010–1018. https://doi.org/10.1002/mds.25776 5. Punsoni M, Friedman JH, Resnick M, et al (2019) Enteric Pathologic Manifestations of Alpha-Synucleinopathies. Appl Immunohistochem Mol Morphol 27:543–548. https://doi.org/10.1097/PAI.0000000000000613 6. Aldecoa I, Navarro-Otano J, Stefanova N, et al (2015) Alpha-synuclein immunoreactivity patterns in the enteric nervous system. Neurosci Lett 602:145–149. https://doi.org/10.1016/j.neulet.2015.07.005 7. Stokholm MG, Danielsen EH, Hamilton-Dutoit SJ, Borghammer P (2016) Pathological α-synuclein in gastrointestinal tissues from prodromal Parkinson disease patients. Ann Neurol 79:940–949. https://doi.org/10.1002/ana.24648 8. Lebouvier T, Neunlist M, Bruley des Varannes S, et al (2010) Colonic biopsies to assess the neuropathology of Parkinson’s disease and its relationship with symptoms. PLoS ONE 5:e12728. https://doi.org/10.1371/journal.pone.0012728 9. Braak H, de Vos RAI, Bohl J, Del Tredici K (2006) Gastric alpha-synuclein immunoreactive inclusions in Meissner’s and Auerbach’s plexuses in cases staged for Parkinson’s disease-related brain pathology. Neurosci Lett 396:67–72. https://doi.org/10.1016/j.neulet.2005.11.012 10. Qualman SJ, Haupt HM, Yang P, Hamilton SR (1984) Esophageal Lewy bodies associated with ganglion cell loss in achalasia. Similarity to Parkinson’s disease. Gastroenterology 87:848–856. https://doi.org/10.1016/0016-5085(84)90079-9 11. Wakabayashi K, Takahashi H, Takeda S, et al (1988) Parkinson’s disease: the presence of Lewy bodies in Auerbach’s and Meissner’s plexuses. Acta Neuropathol 76:217–221. https://doi.org/10.1007/BF00687767 12. Maroteaux L, Campanelli JT, Scheller RH (1988) Synuclein: a neuron-specific protein localized to the nucleus and presynaptic nerve terminal. J Neurosci 8:2804–2815. https://doi.org/10.1523/JNEUROSCI.08-08-02804.1988 13. Burré J, Sharma M, Südhof TC (2018) Cell Biology and Pathophysiology of α-Synuclein. Cold Spring Harb Perspect Med 8(3):a024091. https://doi.org/10.1101/cshperspect.a024091 14. Bartels T, Choi JG, Selkoe DJ (2011) α-Synuclein occurs physiologically as a helically folded tetramer that resists aggregation. Nature 477:107–110. ttps://doi.org/10.1038/nature10324 15. Fauvet B, Mbefo MK, Fares M-B, et al (2012) α-Synuclein in central nervous system and from erythrocytes, mammalian cells, and Escherichia coli exists predominantly as disordered monomer. J Biol Chem 287:15345–15364. https://doi.org/10.1074/jbc.M111.318949 16. Conway KA, Harper JD, Lansbury PT (2000) Fibrils formed in vitro from alpha-synuclein and two mutant forms linked to Parkinson’s disease are typical amyloid. Biochemistry 39:2552–2563. https://doi.org/10.1021/bi991447r 17. Sharrad DF, de Vries E, Brookes SJH (2013) Selective expression of α-synuclein-immunoreactivity in vesicular acetylcholine transporter-immunoreactive axons in the guinea pig rectum and human colon. J Comp Neurol 521:657–676. https://doi.org/10.1002/cne.23198 18. Phillips RJ, Walter GC, Wilder SL, et al (2008) Alpha-synuclein-immunopositive myenteric neurons and vagal preganglionic terminals: autonomic pathway implicated in Parkinson’s disease? Neuroscience 153:733–750. https://doi.org/10.1016/j.neuroscience.2008.02.074 19. Corbillé A-G, Neunlist M, Derkinderen P (2016) Cross-linking for the analysis of α-synuclein in the enteric nervous system. J Neurochem 139:839–847. https://doi.org/10.1111/jnc.13845 20. Böttner M, Fricke T, Müller M, et al (2015) Alpha-synuclein is associated with the synaptic vesicle apparatus in the human and rat enteric nervous system. Brain Res 1614:51–59. https://doi.org/10.1016/j.brainres.2015.04.015 21. Swaminathan M, Fung C, Finkelstein DI, et al (2019) α-Synuclein Regulates Development and Function of Cholinergic Enteric Neurons in the Mouse Colon. Neuroscience 423:76–85. https://doi.org/10.1016/j.neuroscience.2019.10.029 22. Spillantini MG, Schmidt ML, Lee VM, et al (1997) Alpha-synuclein in Lewy bodies. Nature 388:839–840. https://doi.org/10.1038/42166 23. Fujiwara H, Hasegawa M, Dohmae N, et al (2002) alpha-Synuclein is phosphorylated in synucleinopathy lesions. Nat Cell Biol 4:160–164. https://doi.org/10.1038/ncb748 24. Kuusisto E, Parkkinen L, Alafuzoff I (2003) Morphogenesis of Lewy bodies: dissimilar incorporation of alpha-synuclein, ubiquitin, and p62. J Neuropathol Exp Neurol 62:1241–1253. https://doi.org/10.1093/jnen/62.12.1241 25. Beach TG, White CL, Hamilton RL, et al (2008) Evaluation of alpha-synuclein immunohistochemical methods used by invited experts. Acta Neuropathol 116:277–288. https://doi.org/10.1007/s00401-008-0409-8 26. Muntané G, Ferrer I, Martinez-Vicente M (2012) α-synuclein phosphorylation and truncation are normal events in the adult human brain. Neuroscience 200:106–119. https://doi.org/10.1016/j.neuroscience.2011.10.042 27. Anderson JP, Walker DE, Goldstein JM, et al (2006) Phosphorylation of Ser-129 is the dominant pathological modification of alpha-synuclein in familial and sporadic Lewy body disease. J Biol Chem 281:29739–29752. https://doi.org/10.1074/jbc.M600933200 28. Jung BC, Lim Y-J, Bae E-J, et al (2017) Amplification of distinct α-synuclein fibril conformers through protein misfolding cyclic amplification. Exp Mol Med 49:e314. https://doi.org/10.1038/emm.2017.1 29. Corbillé A-G, Preterre C, Rolli-Derkinderen M, et al (2017) Biochemical analysis of α-synuclein extracted from control and Parkinson’s disease colonic biopsies. Neurosci Lett 641:81–86. https://doi.org/10.1016/j.neulet.2017.01.050 30. Fenyi A, Leclair-Visonneau L, Clairembault T, et al (2019) Detection of alpha-synuclein aggregates in gastrointestinal biopsies by protein misfolding cyclic amplification. Neurobiol Dis 129:38–43. https://doi.org/10.1016/j.nbd.2019.05.002 31. Gray MT, Munoz DG, Gray DA, et al (2014) Alpha-synuclein in the appendiceal mucosa of neurologically intact subjects. Mov Disord 29:991–998. https://doi.org/10.1002/mds.25779 32. Killinger BA, Madaj Z, Sikora JW, et al (2018) The vermiform appendix impacts the risk of developing Parkinson’s disease. Sci Transl Med 10(465):earr5280. https://doi.org/10.1126/scitranslmed.aar5280 33. Recasens A, Ulusoy A, Kahle PJ, et al (2018) In vivo models of alpha-synuclein transmission and propagation. Cell Tissue Res 373:183–193. https://doi.org/10.1007/s00441-017-2730-9 34. Dehay B, Vila M, Bezard E, et al (2016) Alpha-synuclein propagation: New insights from animal models. Mov Disord 31:161–168. https://doi.org/10.1002/mds.26370 35. Recasens A, Dehay B, Bové J, et al (2014) Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys. Ann Neurol 75:351–362. https://doi.org/10.1002/ana.24066 36. Recasens A, Carballo-Carbajal I, Parent A, et al (2018) Lack of pathogenic potential of peripheral α-synuclein aggregates from Parkinson’s disease patients. Acta Neuropathol Commun 6:8. https://doi.org/10.1186/s40478-018-0509-1 37. Forno LS, Norville RL (1976) Ultrastructure of Lewy bodies in the stellate ganglion. Acta Neuropathol 34:183–197. https://doi. org/10.1007/BF00688674 38. Del Tredici K, Hawkes CH, Ghebremedhin E, Braak H (2010) Lewy pathology in the submandibular gland of individuals with incidental Lewy body disease and sporadic Parkinson’s disease. Acta Neuropathol 119:703–713. https://doi.org/10.1007/s00401-010-0665-2 39. Greffard S, Verny M, Bonnet A-M, et al (2006) Motor score of the Unified Parkinson Disease Rating Scale as a good predictor of Lewy body-associated neuronal loss in the substantia nigra. Arch Neurol 63:584–588. https://doi.org/10.1001/archneur. 63.4.584 40. Edwards LL, Quigley EM, Pfeiffer RF (1992) Gastrointestinal dysfunction in Parkinson’s disease: frequency and pathophysiology. Neurology 42:726–732. https://doi.org/10.1212/WNL.42.4.726 41. Knowles CH, De Giorgio R, Kapur RP, et al (2009) Gastrointestinal neuromuscular pathology: guidelines for histological techniques and reporting on behalf of the Gastro 2009 International Working Group. Acta Neuropathol 118:271–301. https://doi.org/10.1007/s00401-009-0527-y 42. Annerino DM, Arshad S, Taylor GM, et al (2012) Parkinson’s disease is not associated with gastrointestinal myenteric ganglion neuron loss. Acta Neuropathol 124:665–680. https://doi.org/10.1007/s00401-012-1040-2 43. Giancola F, Torresan F, Repossi R, et al (2017) Downregulation of neuronal vasoactive intestinal polypeptide in Parkinson’s disease and chronic constipation. Neurogastroenterol Motil 29(5):10.1111. https://doi.org/10.1111/nmo.12995 44. Lionnet A, Wade MA, Corbillé A-G, et al (2018) Characterisation of tau in the human and rodent enteric nervous system under physiological conditions and in tauopathy. Acta Neuropathol Commun 6:65. https://doi.org/10.1186/s40478-018-0568-3 45. Guo T, Noble W, Hanger DP (2017) Roles of tau protein in health and disease. Acta Neuropathol 133:665–704. https://doi.org/10.1007/s00401-017-1707-9 46. Dugger BN, Hoffman BR, Scroggins A, et al (2019) Tau immunoreactivity in peripheral tissues of human aging and select tauopathies. Neurosci Lett 696:132–139. https://doi.org/10.1016/j.neulet.2018.12.031 47. Ohlsson B, Englund E (2019) Atrophic Myenteric and Submucosal Neurons Are Observed in Parkinson’s Disease. Parkinsons Dis 2019:7935820. https://doi.org/10.1155/2019/7935820 48. Ozawa T, Shimizu H, Matsui H, et al (2019) Shrinkage of the myenteric neurons of the small intestine in patients with multiple system atrophy. Auton Neurosci 221:102583. https://doi.org/10.1016/j.autneu.2019.102583 49. Rossi M, Candelise N, Baiardi S, et al (2020) Ultrasensitive RT-QuIC assay with high sensitivity and specificity for Lewy body-associated synucleinopathies. Acta Neuropathol 140:49–62. https://doi.org/10.1007/s00401-020-02160-8 50. Arotcarena M-L, Dovero S, Prigent A, et al (2020) Bidirectional gut-to-brain and brain-to-gut propagation of synucleinopathy in non-human primates. Brain 143:1462–1475. https://doi.org/10.1093/brain/awaa096 51. Beach TG, Corbillé A-G, Letournel F, et al (2016) Multicenter Assessment of Immunohistochemical Methods for Pathological Alpha-Synuclein in Sigmoid Colon of Autopsied Parkinson’s Disease and Control Subjects. J Parkinsons Dis 6:761–770. https://doi.org/10.3233/JPD-160888
Copyright: © 2020 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |