|
|
|
Free Neuropathology 1:6 (2020) |
|
Original Paper |
|
Deposits of disease-associated alpha-synuclein may be present in the dura mater in Lewy body disorders: implications for potential inadvertent transmission by surgery |
|
Ellen Gelpi MD1,2, Naomi P. Visanji PhD3, Selma Hönigschnabl MD4, Angelika Reiner MD4, Peter Fischer MD5, Anthony E. Lang MD3, Herbert Budka MD1, Gabor G. Kovacs MD3,6,7 |
|
1 Division of Neuropathology and Neurochemistry, Department of Neurology, Medical University of Vienna, Vienna, Austria |
|
Corresponding author: |
|
Submitted: 09 January 2020 Accepted: 06 February 2020 Published: 12 February 2020 |
|
Keywords: Alpha-synuclein, Dura mater, Lymphatic drainage, Prion, Propagation, Transmission |
|
Abstract Deposition of alpha-synuclein in the brain is a hallmark of Lewy body disorders. Alpha-synuclein has been considered to show prion-like properties. Prion diseases can be transmitted by the transplantation of cadaveric dura mater causing iatrogenic Creutzfeldt-Jakob disease. Recent observations of amyloid-β deposition in dural grafts support the seeding properties of amyloid-β. Here we assessed the presence of alpha-synuclein in dura mater samples as a potential transmissible seed source. We immunostained 32 postmortem dura mater samples; 16 cases with Lewy-body disorder (LBD) showing different pathology stages and 16 non-LBD cases for phosphorylated (Ser129) and disease-associated (5G4) alpha-synuclein. Disease-associated alpha-synuclein aggregates were identified in intradural nerve fibres and associated with a vessel in a single LBD-Braak stage 4 case. We conclude that alpha-synuclein is detectable, although rarely, in dura mater samples in patients with LBD. The risk of potential transmissibility of dural alpha-synuclein deserves assessment by complementary experimental studies. Introduction Neurodegenerative diseases are considered protein-misfolding disorders. As described in prion diseases, the host physiological protein adopts an abnormal conformation (seed), capable of inducing misfolding and aggregation of the neighboring molecules (e.g. for prion protein: the physiological PrPC converting to disease-associated PrPSc, and PrPSc recruiting more PrPC). In prion disease, strong clinical, epidemiological and experimental evidence has demonstrated human and interspecies transmissibility of PrPSc(1). For several other proteins associated with neurodegenerative conditions, such as Amyloid-β (Aβ), Tau, alpha-synuclein, and TAR DNA binding protein (TDP-43), properties similar to those of PrP have been shown predominantly in an experimental setting (2). The potential of Aβ peptides to be transmitted between humans has been a topic of great interest for public health. Young adults with iatrogenic Creutzfeldt-Jakob-Disease (CJD) following cadaveric pituitary-derived growth hormone in childhood were found to have Aβ deposits in the brain parenchyma and blood vessels (3, 5, 6), as well as in iatrogenic CJD after dura grafting (4). Two studies have reported the presence of Aβ deposits in dura mater samples of postmortem brains of patients with Alzheimer’s disease (AD) and other neurodegenerative pathologies (7, 8). Additionally, neurosurgical procedures and dural grafting during childhood or teenage years have been related to increased cerebral amyloid angiopathy (CAA) decades later (9, 10). These findings indicate that dura mater grafts could be also a source of Aβ seeds capable of inducing the aggregation of Aβ in the CNS of the graft recipients. Lewy-body disorders (LBD) and multiple system atrophy (MSA) are characterized by the accumulation of alpha-synuclein. A growing body of experimental evidence supports prion-like propagation in alpha-synucleinopathies (11-13). Observations in a small number of subjects with Parkinson`s disease (PD) treated with fetal mesencephalic dopaminergic grafts in the striatum, who years later were found to have alpha-synuclein positive LBs in grafted neurons, first suggested the potential of alpha-synuclein propagation from host to graft cells (14, 15). While experimental studies have also successfully demonstrated the transmission potential of MSA alpha-synuclein in genetically modified mouse models (16), observations in humans argue against a human-to-human iatrogenic transmission of MSA and PD (17). Given the potential implications for public health, we posit that thorough assessment of the potential transmissibility of all neurodegenerative proteinopathies is essential (18). Therefore, in this study, we assessed human postmortem dura mater samples, some of which have been previously shown to harbor Aβ deposits (7), for the presence of alpha-synuclein aggregates, which may represent seeds with the potential of propagating further synucleinopathy. Material and methods Formalin-fixed and paraffin-embedded postmortem tissue samples of the dura mater from the left temporal and posterior region as well as cross-sections from the area of the superior sagittal sinus and confluence of sinuses (7), were cut into 5-micrometer thick sections. This study was approved by the Ethics Committee of the Medical University of Vienna. Brains were previously evaluated neuropathologically and a final primary diagnosis and concomitant pathologies were recorded. Fixation time of dura mater samples ranged from 3 to 10 years. Immunohistochemistry of the dura mater samples was performed on an automated immunostainer (DAKO Autostainer Link 48, Glostrup, Denmark) using two different anti-alpha-synuclein antibodies: clone 5G4 (1:4000), Roboscreen, Leipzig, Germany; specific for disease-associated forms and the phospho-specific anti-alpha-synuclein phosphorylated at serine 129 (Wako) (1:2000). In addition, we performed immunohistochemical staining to depict the nerve fibres within the connective tissue of the dura mater: anti-neurofilament protein (NFP), clone 2F11 (1:800), DAKO; PGP 9.5 (1:100), DAKO, rabbit polyclonal; anti-tyrosine hydroxylase, clone 1B5 (1:200), Novocastra, Newcastle, UK; and anti-phosphorylated neurofilaments, clone SMI31 (1:5000), former Covance Research Products, Princeton, NJ, USA, and for blood and lymphatic vascular endothelia (anti-CD34, clone QBEnd/10 (1:100), Novocastra; and podoplanin (1:3000), rabbit polyclonal). DAKO Flex system (peroxidase/DAB) was used for visualization of antibody reactions. Samples were assessed blind to the neuropathological diagnosis. The presence or absence of each specific immunoreactivity was recorded. In addition, double immunofluorescence combining PGP 9.5 and 5G4 antibodies and immunofluorescence staining with NFP and 5G4 on adjacent tissue sections was performed in one selected case. Results We analyzed postmortem dura mater samples from 32 donors (50% females) aged between 84 and 87 years (mean 84.9) from the Vienna Transdanube Aging (VITA) Study (19).
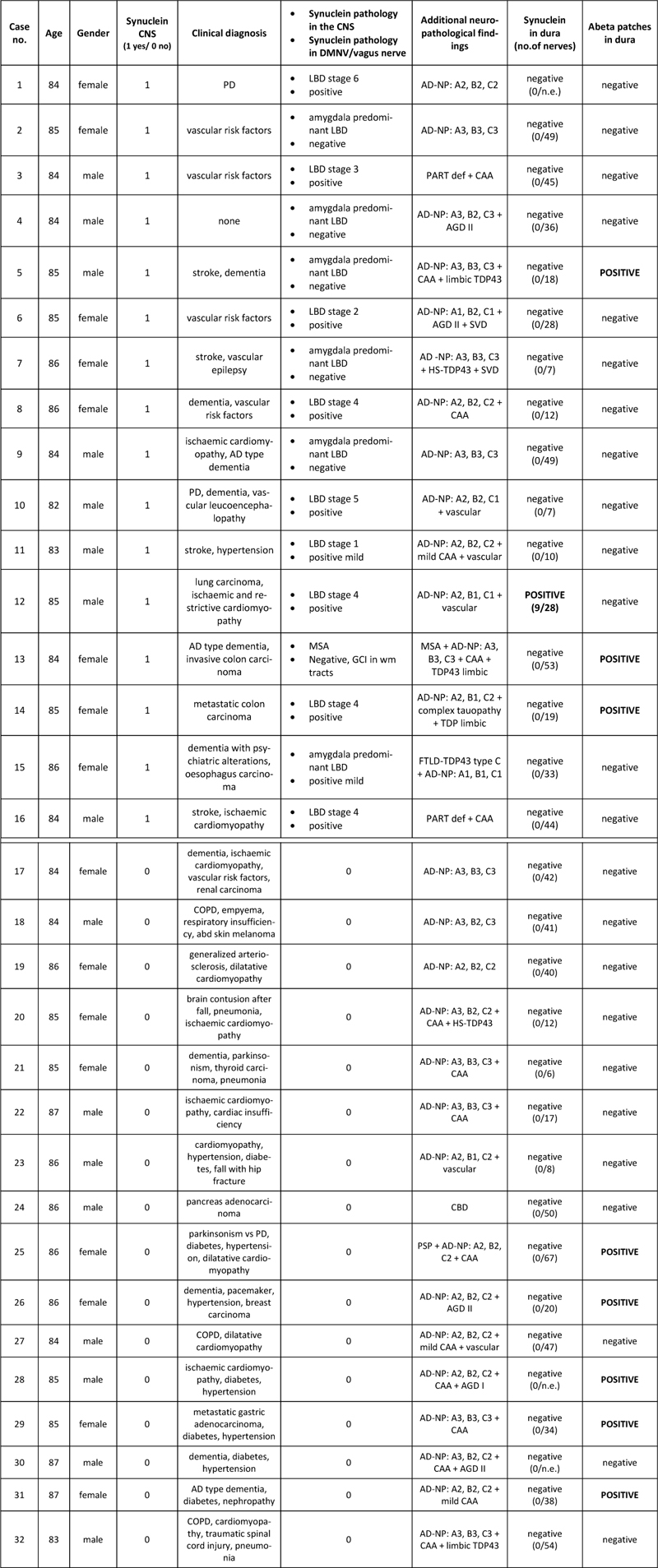
Table 1: Demographic and neuropathological features of the study cohort Abbreviations: AD: Alzheimer’s disease; PD: Parkinson’s disease; COPD: chronic obstructive pulmonary disease; AD-NP: Alzheimer’s disease neuropathological changes; CAA: cerebral amyloid angiopathy; AgD: argyrophilic grain disease; HS: hippocampal sclerosis; PART: primary age-related tauopathy; SVD: small vessel disease; CBD: corticobasal degeneration. n.a.: not available; n.e.: not evaluable due to unsteady immunoreactivity These cases were analyzed in a previous study for the presence of Aβ deposits Neuropathological diagnoses included 16 cases with alpha-synuclein pathology (Table 1; six cases with amygdala predominant Lewy body pathology, one MSA case, single cases of Braak stage 1, stage 2, stage 3, stage 5 and stage 6, and four cases with Braak stage 4 of PD-related pathology) (20); and 16 cases without LB pathology but other neuropathological conditions. Nerve fibers were detected mainly by NFP immunohistochemistry in all cases in variable distribution: large nerves surrounding large vessels (Fig. 1A, B) or smaller nerve branches in between both dura layers (Fig. 1C, D). ). These patterns were also observed by immunofluorescence (Fig. 2). No immunoreactivity for tyrosine-hydroxylase could be detected. CD34 and podoplanin also gave negative results.
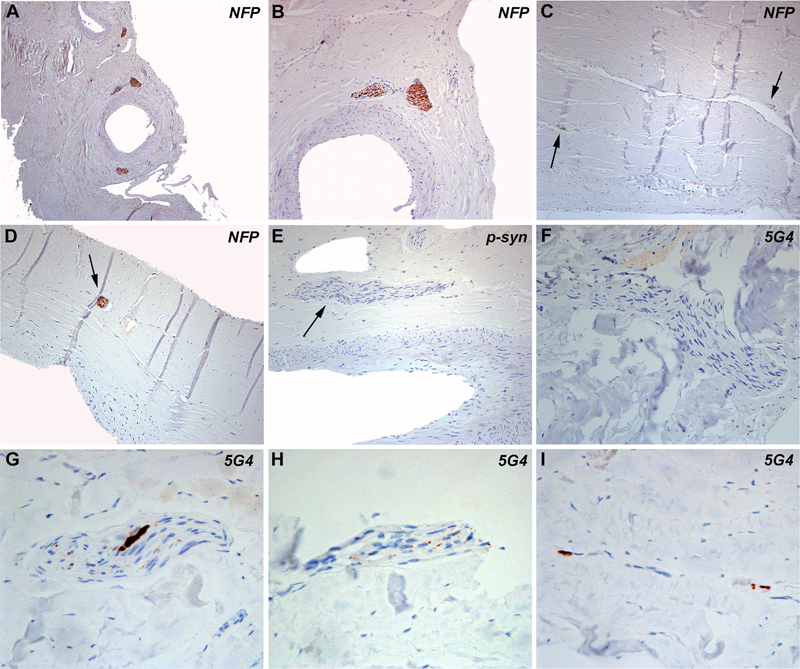
Figure 1 A-D: Histologic appearance of dura mater consisting of two lamellae of connective tissue. Within the connective tissue there are large and small vessels, which are surrounded by nerve fibres (A-D, neurofilament immunohistochemistry). Nerve fibres are also identified between the lamellae and within the connective tissue (arrows). E-I: Immunohistochemistry for phosphorylated alpha-synuclein at serine 129 and for oligomeric forms with the 5G4 antibody did not show abnormal alpha-synuclein aggregates in most of the studied cases (Fig. 1E, F). In one case (case #12 table 1), coarse (G) and punctate (H) aggregates within nerve fibres and perivascular (I) aggregates were identified with the 5G4 antibody. Detailed immunohistochemical analysis of dura samples identified pathological alpha-synuclein aggregates with the 5G4 antibody in one case. These aggregates consisted of small punctate deposits along nerve fibres (Fig. 1H, 2B, D), coarse aggregates (Fig. 1G, 2F) and perivascular deposits (Fig. 1I, 2A). We observed a variable staining intensity of axons among the different cases and also within the same case, which was not related to the presence/absence of alpha-synuclein. This variability might have been related to the fixation time or tissue quality, among other potential variables, so we could not reliably assess axonal preservation in the patient with alpha-synuclein aggregates. We found no tortuous axonal profiles or axonal swellings. This individual was an 85-year-old male with coronary disease with severe and generalized atherosclerosis, an old myocardial infarction, as well as pulmonary adenocarcinoma. No parkinsonism or dementia were clinically reported. Neuropathological examination revealed a vascular encephalopathy and Braak stage 4 Lewy-body pathology, with only mild neurofibrillary pathology in the limbic system and mild neuritic plaque / Aβ pathology (Table 1, case 12). In the remaining cases included in our study, with or without CNS Lewy body pathology, there was a complete lack of alpha-synuclein immunoreactivity in dural samples applying both antibodies.
Figure 2: Immunofluorescence images A, B: Double immunofluorescence for PGP 9.5 depicts delicate perivascular fibres (red) which focally also accumulate alpha-synuclein (5G4, green). Other regions show nerve fascicles with variable density of axonal profiles: some have abundant axons (B, PGP 9.5, red) and show only tiny alpha-synuclein accumulations (5G4, green, white arrow). C-F: In adjacent tissue sections some nerve fascicles have only few axons (C, NFP) while others show an intermediate density of axons (E, NFP red) with relatively abundant and coarse alpha-synuclein aggregates (F, 5G4). Discussion In this study, we found alpha-synuclein aggregates in dura mater samples from one patient with Lewy-body pathology (Braak stage 4) (20). Alpha-synuclein aggregates were not identified in the dura mater of 31 other cases with different stages of LBD or in other neurodegenerative conditions. These findings might have implications for public health as well as for the understanding of alpha-synuclein propagation. Concerning public health, individuals with symptomatic or prodromal alpha-synucleinopathies may undergo neurosurgical interventions contacting the dura mater (for example, the widespread use of deep brain stimulation in PD). While there is still no epidemiological evidence of human-to-human transmission of alpha-synuclein (17), the application of appropriate prevention measures will be increasingly required if experimental studies strengthen evidence of the potential transmissibility of synucleinopathies. This further emphasizes the need for effective biomarkers for the early diagnosis of alpha-synuclein related diseases (21-24). Furthermore, there is a clear need for the detailed mapping of tissues that may harbor potential seeds. Indeed, with respect to the present findings, since dura mater transplants are in direct contact with brain tissues, they might provide an increased potential to propagate seeds to the recipient’s CNS. Concerning protein propagation, the dura mater has been a source of disease transmission in human prion disease (1) and is also a source for Aβ seeds (9, 10). Recent studies support the notion that these seeds are able to propagate pathological proteins, even without inducing the classical Alzheimer´s phenotype. However, they can lead to unexpectedly increased frequency of cerebral amyloid angiopathy and associated brain hemorrhages (9, 10). Propagation of human alpha-synuclein has been considered since the detection of alpha-synuclein aggregates in grafted fetal mesencephalic neurons in PD patients (14, 15). This has subsequently been demonstrated in experimental studies (13) including inoculation of human substantia nigra extracts containing Lewy bodies into wild-type mice and macaque monkeys (26). In contrast, the inoculation of extracts of peripheral alpha-synuclein from the autonomic sympathetic stellate ganglion did not induce pathological conversion of endogenous alpha-synuclein, alpha-synuclein propagation, or neurodegeneration in mice (27). Thus, central and peripheral nervous system-related alpha-synuclein might have different propagation potential and therefore different transmissibility. The dura mater is thoroughly innervated by afferent fibres arising from the ipsilateral trigeminal ganglion and nerve (including its ophthalmic, maxillary and mandibular divisions) and by sympathetic fibres arising from the ipsilateral superior cervical ganglion. The posterior cranial fossa dura mater receives sensory meningeal branches from the vagus and glossopharyngeal nerves. The sympathetic ganglion is upstream from the stellate ganglion, which has been shown to harbor prominent alpha-synuclein or Lewy-type pathology in postmortem studies of patients with PD and dementia with Lewy bodies (23, 24), along with the dorsal motor nucleus of the vagal nerve in the CNS (20). Therefore, it might be reasonable to consider that these sympathetic and parasympathetic fibres innervating the dura mater might also accumulate alpha-synuclein. Although in that particular case where we found alpha-synuclein in the dura mater, we did not have the opportunity to stain the trigeminal ganglion or nerve nor the stellate ganglion, we have previously evaluated the trigeminus nerve using immunostaining for alpha-synuclein. In that study we found alpha-synuclein immunoreactivity in two out of four PD/LBD cases where a total of six trigeminal nerves (in two cases both sides available) were examined (28). Alternatively, we have previously reported that the presence of lymphatic vessels in dura mater might be a drainage route for molecules from the brain parenchyma in neurodegenerative disorders, including alpha-synuclein in LBD (25). Although we found discrete perivascular alpha-synuclein aggregates in dura mater samples, we were not able to confirm whether these represent lymphatic vessels, possibly due to the prolonged fixation time of this dura mater sample (<6 years), and therefore this question deserves further study. The present study has some limitations. First, as mentioned above, the dura mater samples were obtained after a prolonged fixation time, which may influence immunohistochemical results, not only for CD34, podoplanin and TH, but for phospho-synuclein as well (29). Therefore, the interpretation concerning the perivascular association of alpha-synuclein aggregates needs some caution. We could also not reliably assess axonal preservation in the case harboring alpha-synuclein aggregates. The prolonged fixation also does not allow complementary molecular/biochemical studies. While the identification of characteristic pathological aggregates of alpha-synuclein with only one antibody is limiting, the 5G4 antibody has shown to be a robust marker of early alpha-synuclein pathology, even in brains with prolonged fixation time (30). Second, the level of pathological forms of alpha-synuclein detectable by conventional immunohistochemistry may be low. Moreover, the analysis of the supratentorial, rather than the infratentorial dura could also have been a limiting factor due to differences in their innervation; however, the supratentorial dura mater also receives sympathetic and parasympathetic innervation, and thus could also be exposed to transport of misfolded proteins. The negative findings in the other LBD cases of the series might reflect a sampling limitation, as it was not feasible to examine the whole dura mater. Alternatively, the lack of staining could also reflect the diverse innervation of the dura and different pathogenic capacities of central and peripheral alpha-synuclein, or it could also indicate that the dura mater is not a primary vulnerable region for alpha-synuclein aggregation and could be considered to be a low risk tissue for the accumulation of potentially transmissible seeds. It must be emphasized that this study raises a potentially important issue but is unable to provide a conclusion on the risk of transmission of alpha-synuclein via dura mater grafts or neurosurgical procedures. This observation would ideally be complemented by experimental studies using prospectively collected freshly harvested, short fixed and frozen dura samples, including a larger cohort of patients of different age groups. Conclusion Disease-associated alpha-synuclein is detectable by conventional immunohistochemistry in postmortem supratentorial dura mater, here in a single patient with Lewy-body-type alpha-synucleinopathy in the CNS. This suggests propagation of alpha-synuclein from the brain and may indicate that dura mater could be a potential risk tissue for inadvertent transmission of alpha-synuclein by dura mater grafts or surgery. Further experimental and biochemical studies are needed to assess the risk of alpha-synuclein transmission via dura mater and its relationship with human disease. Acknowledgements We are grateful to Susi Schmid, Irene Ebner, Gerda Ricken and Sigrid Klotz from the Division of Neuropathology and Neurochemistry, Department of Neurology, Medical University of Vienna, for excellent technical support. We are indebted to brain donors and their family of the Vienna Transdanube Study (VITA) for generous brain and sample donation for research. This work has been supported by the Bishop Karl Golser Award to GGK. References 1. Brown P, Brandel JP, Sato T, et al. Iatrogenic Creutzfeldt-Jakob disease, final assessment. Emerg Infect Dis. 2012; 18:901-907. 2. Duyckaerts C, Clavaguera F, Potier MC. The prion-like propagation hypothesis in Alzheimer's and Parkinson's disease. Curr Opin Neurol. 2019; 32:266-271. 3. Jaunmuktane Z, Mead S, Ellis M, et al. Evidence for human transmission of amyloid-β pathology and cerebral amyloid angiopathy. Nature. 2015; 525:247-250. 4. Frontzek K, Lutz MI, Aguzzi A, et al. Amyloid-β pathology and cerebral amyloid angiopathy are frequent in iatrogenic Creutzfeldt-Jakob disease after dural grafting. Swiss Med Wkly. 2016; 146:w14287. 5. Silvia A. Purro, Mark A. Farrow, Jacqueline Linehan, et al. Transmission of amyloid-β protein pathology from cadaveric pituitary growth hormone. Nature. 2018; 564:415-419. 6. Burwinkel M, Lutzenberger M, Heppner FL, et al. Intravenous injection of beta-amyloid seeds promotes cerebral amyloid angiopathy (CAA). Acta Neuropathol Commun. 2018; 6:23. 7. Kovacs GG, Lutz MI, Ricken G, et al. Dura mater is a potential source of Aβ seeds. Acta Neuropathol. 2016; 131:911-923. 8. Hamaguchi T, Taniguchi Y, Sakai K, et al. Significant association of cadaveric dura mater grafting with subpial Aβ deposition and meningeal amyloid angiopathy. Acta Neuropathol. 2016; 132:313-315. 9. Jaunmuktane Z, Quaegebeur A, Taipa R, et al. Evidence of amyloid-β cerebral amyloid angiopathy transmission through neurosurgery. Acta Neuropathol. 2018; 135:671-679. 10. Banerjee G, Adams ME, Jaunmuktane Z, et al. Early onset cerebral amyloid angiopathy following childhood exposure to cadaveric dura. Ann Neurol. 2019; 85:284-290. 11. Recasens A, Dehay B, Bové J, et al. Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys. Ann Neurol. 2014; 75:351-362. 12. Karpowicz RJ Jr, Trojanowski JQ, Lee VM. Transmission of α-synuclein seeds in neurodegenerative disease: recent developments. Lab Invest. 2019; 99:971-981. 13. Desplats P, Lee HJ, Bae EJ, et al. Inclusion formation and neuronal cell death through neuron-to-neuron transmission of alpha-synuclein. Proc Natl Acad Sci U S A. 2009; 106:13010-13015. 14. Kordower JH, Chu Y, Hauser RA, et al. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson's disease. Nat Med. 2008; 14:504-506. 15. Li JY, Englund E, Holton JL, et al. Lewy bodies in grafted neurons in subjects with Parkinson's disease suggest host-to-graft disease propagation. Nat Med. 2008; 14:501-503. 16. Woerman AL, Oehler A, Kazmi SA, et al. Multiple system atrophy prions retain strain specificity after serial propagation in two different Tg(SNCA*A53T) mouse lines. Acta Neuropathol. 2019; 137:437-454. 17. De Pablo-Fernandez E, Cerdán Santacruz D, Warner TT, et al. No evidence of iatrogenic human transmission in autopsy confirmed multiple system atrophy. Mov Disord. 2018; 33:1183-1184. 18. De Sousa PA, Ritchie D, Green A, et al. Renewed assessment of the risk of emergent advanced cell therapies to transmit neuroproteinopathies Acta Neuropathol. 2018; 137:363-377. 19. Fischer P, Jungwirth S, Krampla W, et al. Vienna Transdanube Aging "VITA": study design, recruitment strategies and level of participation. J Neural Transm Suppl. 2002; 62:105-116. 20. Braak H, Del Tredici K, Rüb U, et al. Staging of brain pathology related to sporadic Parkinson's disease. Neurobiol Aging. 2003; 24:197-211. 21. Vilas D, Iranzo A, Tolosa E, et al. Assessment of α-synuclein in submandibular glands of patients with idiopathic rapid-eye-movement sleep behaviour disorder: a case-control study. Lancet Neurol. 2016; 15:708-718. 22. Tolosa E, Gaig C, Santamaría J, et al. Diagnosis and the premotor phase of Parkinson disease. Neurology. 2009; 72(7 Suppl):S12-20. 23. Gelpi E, Navarro-Otano J, Tolosa E, et al. Multiple organ involvement by alpha-synuclein pathology in Lewy body disorders. Mov Disord. 2014; 29:1010-1018. 24. Beach TG, Adler CH, Sue LI, et al; Arizona Parkinson's Disease Consortium. Multi-organ distribution of phosphorylated alpha-synuclein histopathology in subjects with Lewy body disorders. Acta Neuropathol. 2010; 119:689-702. 25. Visanji NP, Lang AE, Munoz DG. Lymphatic vasculature in human dorsal superior sagittal sinus: implications for neurodegenerative proteinopathies. Neurosci Lett. 2018; 665:18-21. 26. Recasens A, Dehay B, Bové J, et al. Lewy body extracts from Parkinson disease brains trigger α-synuclein pathology and neurodegeneration in mice and monkeys. Ann Neurol. 2014; 75:351-362. 27. Recasens A, Carballo-Carbajal I, Parent A, et al. Lack of pathogenic potential of peripheral α-synuclein aggregates from Parkinson's disease patients. Acta Neuropathol Commun. 2018; 6:8. 28 Kovacs GG, Breydo L, Green R, et al. Intracellular processing of disease-associated α-synuclein in the human brain suggests prion-like cell-to-cell spread. Neurobiol Dis. 2014; 69:76-92. 29. Pikkarainen M, Martikainen P, Alafuzoff I. The effect of prolonged fixation time on immunohistochemical staining of common neurodegenerative disease markers. J Neuropathol Exp Neurol. 2010; 69:40-52. 30. Kovacs GG, Wagner U, Dumont B, et al. An antibody with high reactivity for disease-associated α-synuclein reveals extensive brain pathology. Acta Neuropathol. 2012; 124:37-50.
Copyright: © 2020 The author(s). This is an open access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited, a link to the Creative Commons license is provided, and any changes are indicated. The Creative Commons Public Domain Dedication waiver (https://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. |