Main Research Topics
- Gene architecture and expression
- Splicing errors and their evolutionary significance
- Organismal responses to environmental stress
- Transgenerational epigenetic inheritance

Genes are fascinating. “Genes influence what we look like on the outside and how we work on the inside ” (Virtual Genetics Education Center, University of Leicester). Humans have an estimated number of ~20,000 protein-coding genes, a single-celled organism like the ciliate Paramecium ~40,000. Different gene numbers are estimated for different eukaryotes. Where do genes come from? And why does gene number differ between species?

Eukaryotic genes (such as those in human and Paramecium) contain intervening noncoding DNA sequences known as spliceosomal introns. Like exons, which regulate or contain the instructions for protein synthesis, introns are transcribed into precursor mRNA (pre-mRNA). Unlike exons, introns are immediately removed from pre-mRNAs through a process known as RNA splicing. Where do introns come from? And why do cells eliminate introns immediately after transcribing them? Modern economic theories would consider this an unacceptable waste of energy. Where is the catch?
In humans and many other vertebrates, introns are very large (amongst the largest across eukaryotes and much larger than their contiguous exons). In contrast, in microbial species such as Paramecium, introns (but not exons) are extremely small. What are the reasons behind these differences in exon-intron structures? And why do cells even have introns at all, given that many diseases caused by mutations can be linked to RNA splicing? These and many other questions remain unanswered. They have attracted and continue to attract the attention of thousands of scholars from various fields of research. Members of the Catania lab are amongst them.

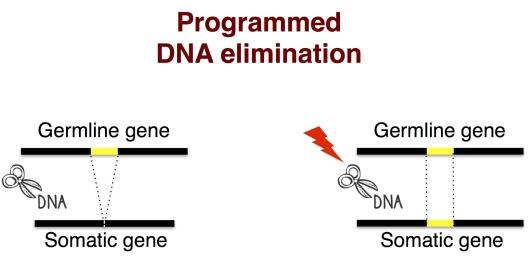
In many eukaryotes (such as human and Paramecium) splicing occurs also at the DNA level, upon sexual reproduction. Germline sequences may be eliminated to regenerate a functional somatic genome (e.g., during the maturation of lymphocytes in humans) through a process known as Programmed DNA Elimination or DNA splicing. As a result of suboptimal splicing signals, splicing may fail partially or entirely to remove target sequences. When defective, splicing may give rise to diseases. At the same time, its inaccuracies generates distinct isoforms that, by affecting the phenotype on occasion, can become substrates of natural selection. How (often) do splicing-derived adaptive variants emerge? Do changes in the environment affect splicing accuracy and the degree to which spliced variants can be inherited across generations? Splicing events are influenced by patterns of epigenetic modifications, which are themselves susceptible to environmental variations. It follows that both the genomic and the surrounding environment must be considered to gain a complete understanding of the role that splicing has in the emergence and establishment of functional genetic novelties.
Gene architecture may vary dramatically across eukaryotic species. These variations are due to phylogenetic differences in population-genetic and intracellular environments. While the role of the population-genetic forces in shaping eukaryotic gene features has been given some consideration, the contribution of the intracellular processes remains largely unexplored. In beginning to fill this knowledge gap, we found that the exon-intron structure of Drosophila genes is nonrandom and reflects the mode of operation of co-transcriptional processes (Lepennetier and Catania, G3, 2016). These and other properties of eukaryotic genes (Lepennetier and Catania, G3, 2017) are consistent with the hitherto uncharted hypothesis that interacting mRNA-associated processes such as capping, RNA splicing, cleavage/polyadenylation, etc. can affect the evolution of eukaryotic gene architecture (Catania and Lynch, Bioessays 2013; Catania, Int. J. Biochem. and Cell Biol. 2017).

Eukaryotic genomes are highly dynamic units. They change even within an organism’s lifetime. For example, during the development of vertebrate embryos, portions of the germline DNA are excised to enable the formation of functional antibody genes in specialized somatic cells called lymphocytes. Similar DNA-level splicing events occur in Paramecium at the genome-wide level. Like all ciliates, Paramecium contains two nuclei. A somatic genome is housed in the macronucleus and a germline genome in the micronucleus. The germline genome contains thousands of Internal Eliminated Sequences (or IESs), i.e., short DNA sequences that reside within and outside genes. After sexual reproduction, the germline DNA regenerates the somatic DNA. During this developmental process the germline DNA is fragmented, amplified and thousands of IESs are excised to enable the formation of functional somatic genes.
In Paramecium, IES excision has been found to be imperfect, as the removal of germline DNA sequences from the newly developed polyploid somatic genome is often incomplete (Duret et al., Genome Research, 2008). Our own analyses support these findings and further suggest that a subset of IESs is differentially retained between P. aurelia species (Catania et al., Genome Biology and Evolution, 2013).
We have recently introduced a measure for the quality of cis-acting DNA-level splicing signals, termed Cin-score (Ferro et al, Nucleic Acids Research, 2015). The Cin-score is both correlated with IES length and shaped by natural selection. Increased Cin-scores may enhance the recognition/excision of large IESs by perhaps assisting in and/or stabilizing the formation of double-stranded DNA loops.
Currently we are investigating the effect of environmental variations on the emergence of distinct (RNA and DNA) splicing variants. We want to assess the effect of environmental—temperature—variations on the emergence and the fate of new splicing isoforms over subsequent sexual generations of Paramecium. This exploratory study 1) should provide an adequate sense of the effect that temperature variations have on IES excision and RNA splicing, and 2) will permit us to monitor changes in the relative fraction of imperfect IES excisions over time. Our preliminary work suggests that temperature variations perturb the efficiency of the DNA excision machinery and that Cin-scores affect significantly the likelihood for a DNA region of being inaccurately excised.
The underlying physiological mechanisms that link reproduction to the ability to survive environmental stress are not yet fully understood. We are currently addressing this question. We are exploring how the relationship between these traits varies with age, population density, and degree of caloric restriction using Paramecium as model organism. Work in progress.