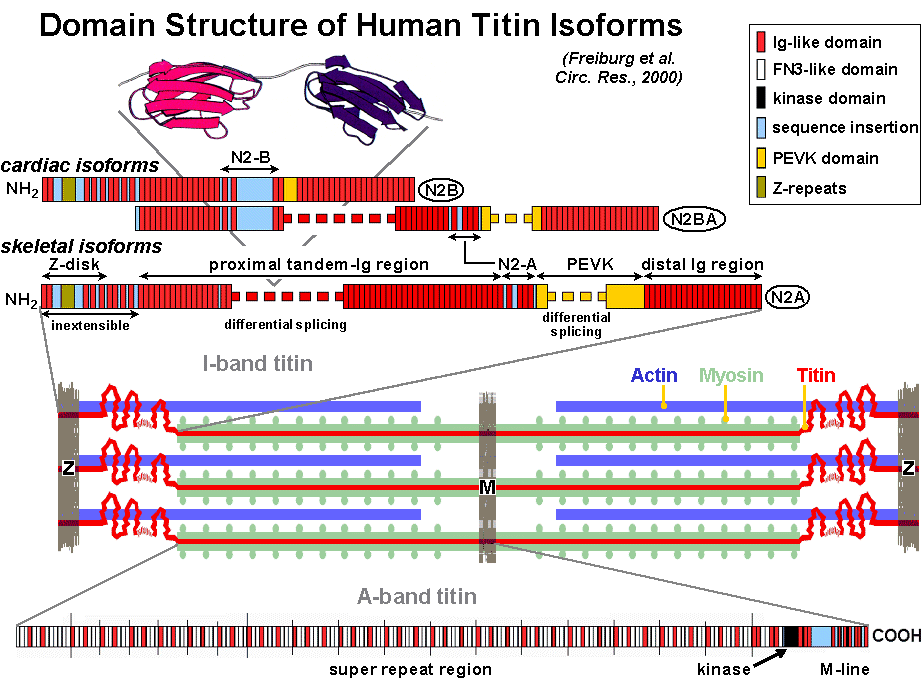
Titin is a high-molecular weight protein (3.0-3.8 MDa) in the sarcomeres of vertebrate striated muscles. A plethora of titin isoforms is expressed from the copy of a single titin gene (on chromosome 2 in mouse and humans). A current view of the arrangement of titin in the sarcomere and titin's domain architecture is shown in Fig 1. Titin's role in muscle structure and function is multifaceted. The polypeptide (1) provides a structural framework through association with other proteins of the sarcomere, (2) keeps the thick filament centered in the sarcomere during activation, (3) functions as a molecular spring in muscle sarcomeres, and (4) is involved in myocyte signaling, associates with signaling molecules and probably has a central place in the stress-sensing machinery of the muscle cell. Currently more than 15 sarcomeric and non-sarcomeric proteins are known to interact with titin. Elucidation of titin's primary structure revealed that the polypeptide contains, near its COOH-terminal end (M-line), a kinase domain (Fig. 1), which may be active during muscle development. Titin also contains a number of phosphorylation sites, in the M-line, Z-disk and I-band, which may be important for the function of titin in signal transduction.
Elastic region of titin. A particularly important property of titin filaments is their ability to act as molecular springs providing relaxed myofibrils with elasticity. Although single titin polypeptides span the entire half of a sarcomere, only a molecular segment confined to the I-band is functionally elastic (Fig. 1). This segment consists of two main, structurally distinct, regions. One region is a unique sequence rich in proline (P), glutamate (E), valine (V) and lysine (K) residues, termed the PEVK segment, which has no obvious secondary or tertiary structure. The PEVK domain is flanked on either side by a region made up of serially linked immunoglobulin-like (Ig) domains--modules that fold into seven-stranded beta-barrels. Both regions are expressed in muscle-type specific length isoforms. Skeletal-muscle sarcomeres contain a so-called N2-A isoform, whose length varies in different tissue types. Cardiac cells co-express an N2-B isoform and N2-BA isoforms, the latter of which contain structural elements from both the N2-A and N2-B isoforms and vary in length in their proximal Ig segments and PEVK-domain. Cardiac titin also contains a unique region termed "N2-B" (Fig. 1). We demonstrated that the Ig-domain regions confer extensibility to the sarcomere mainly at low stretch, the PEVK domain predominantly at higher stretch. In cardiac muscle also the N2-B unique sequence extens at high stretch forces. The elastic region of titin interacts with several other proteins, e.g., actin and the chaperone alpha-B-crystallin, but other proteins important for myocyte signaling and turnover as well. Weak interactions of titin's PEVK-domain with the thin filament give rise to a viscous force component during quick length changes of muscle.
Molecular basis of titin elasticity. State-of-the-art techniques have been used to explore titin elasticity at the single molecule level. Laser trapping and atomic force microscopy (AFM) studies on native or engineered titin molecules or titin domains have revealed a nonlinear force response upon stretching, which was explained in terms of the entropic-chain characteristics of the polypeptide. An entropic chain undergoes thermally-induced bending movements that tend to shorten its end-to-end length (Fig. 2). Stretching such a chain reduces its conformational entropy and thus, requires an external force. In the mechanical studies on isolated proteins, titin's force-extension relation could be well fitted with the wormlike chain (WLC) model of entropic elasticity. A WLC is a deformable rod whose bending rigidity is characterized by its persistence length, a distance within which the orientations of the chain are correlated. Rigid polymers have a long persistence length (relative to their contour length) and their straightening requires little external force, while the opposite is true for more flexible polymers. In titin, the Ig-domain regions represent WLC elements with relatively long persistence length. In addition, these regions appear to be capable of adjusting their contour lengths by unfolding a few of their Ig modules. Ig-domain unfolding has been readily shown in AFM stretch experiments on single recombinant polyproteins containing only a small number of Ig modules. The PEVK domain and the N2-B unique sequence are entropic springs with a relatively short persistence length, which explains why these regions extend after the Ig-domain segments. Currently it is debated whether titin's unique sequences also have an "enthalpic" component of elasticity, brought about e.g. by electrostatic or hydrophobic interactions.

Titin and active force generation. The titin filament could play some active role in contraction. The elastic section of titin exerts a retractive force, which tends to shorten the sarcomere (in cardiac muscle at end diastole, this force can be appreciable); the force is effectively contractile. It has been proposed that titin could stiffen upon activation--already before sarcomeric stiffness increases when myosin cross-bridges attach strongly to actin--perhaps due to the rise in intracellular [Ca2+]. Titin's "contractile" force would thus increase. Indeed, calcium binds to titin at the PEVK domain. Thus, the possibility exists that Ca2+ affects titin stiffness. Recently, we found that the PEVK domain of cardiac titin interacts with actin filaments in a Ca2+-regulated manner, thereby modulating thin-thick filament interaction. It is likely that titin plays other important roles for active contractile functions in cardiac muscle, e.g., through an effect on the length-dependent contractility of the heart (the so-called Frank-Starling mechanism).
Cardiac titin and human heart disease. Human-heart sarcomeres coexpress a stiffer N2B titin and more compliant N2BA titin isoforms. The ratio between N2BA and N2B titin is about 30:70 in normal human heart, but can increase to approximately 50:50 in failing hearts from coronary artery disease (CAD) patients. The titin-isoform shift leads to decreased titin-derived stiffness of the cardiomyofibrils, although whole hearts usually are globally stiffened due to increased fibrosis. Thus, titin can be modified in response to chronic human heart disease. Higher (sometimes perhaps lower) than normal or unchanged N2BA:N2B titin ratios may be found under different disease conditions.
Cardiac titin in heart development. Before
birth the hearts of
rats, mice, pigs, and other species, express very long, compliant,
embryonic/fetal titin
isoforms of the N2BA-type, which can be as large as 3.7 MDa. The
postnatal heart contains
much shorter, stiffer, titin isoforms, including the N2B-isoform.
Skeletal titin isoforms and muscle fiber type. The isoform sizes of titin in different skeletal muscles of rabbits vary between ~3.3 and 3.7 MDa. Titin size correlates modestly with the fiber type and myosin heavy chain isoform in that slower muscles tend to express larger titins than faster muscles (Prado et al., 2005). Titin stiffness adds substantially to the passive stiffness of the whole muscle but titin's contribution to total passive stiffness, relative to that of extracellular elements (collagen), varies greatly among different muscles.
Further reading (related original publications from our laboratory):
Bullard B., C. Ferguson, A. Minajeva, M.C. Leake, M. Gautel, D. Labeit, L. Ding, S. Labeit, J. Horwitz, K. Leonard & W.A. Linke (2004) Association of the chaperone alpha-B-crystallin with titin in heart muscle. J. Biol. Chem. 279:7917-7924. free pdf from publisher
Kulke M., S. Fujita-Becker, E. Rostkova, C. Neagoe, D. Labeit, D.J. Manstein, M. Gautel & W.A. Linke (2001) Interaction between PEVK-titin and actin filaments: origin of a viscous force component in cardiac myofibrils. Circ. Res. 89:874-881. pdf available free from publisher, plus online data supplement
Kulke M., C. Neagoe, B. Kolmerer, A. Minajeva, H. Hinssen, B. Bullard & W.A. Linke (2001) Kettin, a major source of myofibrillar stiffness in Drosophila indirect flight muscle. J. Cell Biol. 154:1045-1057. News coverage in J. Cell Biol. and pdf available free from publisher.
Li H., W.A. Linke, A.F. Oberhauser, M. Carrion-Vazquez, J.B. Kerkvliet, H. Lu, P.E. Marszalek & J.M. Fernandez (2002) Reverse engineering of the giant muscle protein titin. Nature. 418:998-1002. free pdf available here plus online supplement, and News coverage in J. Cell Biol.
Linke W.A., M. Ivemeyer, N. Olivieri, B. Kolmerer, J.C. Rüegg & S. Labeit (1996) Towards a molecular understanding of the elasticity of titin. J. Mol. Biol. 261:62-71. free pdf here
Linke W.A., M. Ivemeyer, P. Mundel, M.R. Stockmeier & B. Kolmerer (1998) Nature of PEVK-titin elasticity in skeletal muscle. Proc. Natl. Acad. Sci. USA. 95:8052-8057. pdf available free from publisher.
Linke W.A., D.E. Rudy, T. Centner, M. Gautel, C. Witt, S. Labeit
& C.C. Gregorio
(1999) I-band titin in cardiac muscle is a three-element molecular
spring and is critical
for maintaining thin filament structure. J. Cell Biol. 146:631-644. pdf
available free from
publisher.
Makarenko, I., C.A. Opitz, M.C. Leake, C. Neagoe, M. Kulke, J.K. Gwathmey, F. del Monte, R.J. Hajjar & W.A. Linke (2004) Passive stiffness changes caused by upregulation of compliant titin isoforms in human dilated cardiomyopathy hearts. Circ. Res. 95:708-716. free pdf from publisher and free online data supplement
Minajeva, A., C. Neagoe, M. Kulke & W.A. Linke (2002) Titin-based contribution to shortening velocity of rabbit skeletal myofibrils. J. Physiol. 540:177-188. pdf available free from publisher.
Minajeva A., M. Kulke, J.M. Fernandez & W.A. Linke (2001) Unfolding of titin domains explains the viscoelastic behavior of skeletal myofibrils. Biophys. J. 80:1442-1451. pdf available free from publisher.
Neagoe C., M. Kulke, F. del Monte, J.K. Gwathmey, P.P. de Tombe, R.J. Hajjar & W.A. Linke (2002) Titin isoform switch in ischemic human heart disease. Circulation. 106:1333-1341. free pdf from publisher
Opitz,
Opitz, C.A., M.C. Leake, I. Makarenko, V. Benes & W.A. Linke (2004) Developmentally regulated switching of titin size alters myofibrillar stiffness in the perinatal heart. Circ. Res. 94:967-975. free pdf from publisher , online data supplement , and Editorial
Prado, L., I. Makarenko, C. Andresen, M. Krüger, C.A. Opitz & W.A. Linke (2005) Isoform diversity of giant proteins in relation to passive and active contractile properties of rabbit skeletal muscles. J. Gen. Physiol. 126:461-480. free pdf available here (! Large file: 11.5 MB !)
Linke,
W.A.
(2007) Sense and stretchability: The role of titin and
titin-associated proteins in myocardial stress-sensing and mechanical
dysfunction. Cardiovasc. Res. April 06, 2007 Epub ahead of print. doi:
10.1016/j.cardiores.2007.03.029. free pdf
here
Labeit, S., B. Kolmerer & W.A. Linke (1997) The giant protein titin: Emerging roles in physiology and pathophysiology. Circ. Res. 80:290-294. free full text from publisher
Linke, W.A. (2000) Stretching molecular springs: Elasticity of titin filaments in vertebrate striated muscle. Histol. Histopathol. 15:799-811. free pdf available here.
Linke, W.A. & H. Granzier (1998) A spring tale: New facts on titin elasticity. Biophys. J. 75:2613-2614. pdf available free from publisher.
Neagoe,